
正常人红细胞的生成包括:造血干细胞阶段、红系祖细胞阶段、红系前体细胞(原红细胞至晚红细胞)的增殖与分化阶段、网织红细胞的增殖及成熟过程,以及网织红细胞向外周血释放成熟为红细胞的过程。
造血干细胞阶段
目前已知,造血干细胞主要存在于骨髓、脾、肝等造血组织内。也有少量循环于外周血中。
造血干细胞在体内的数量极少,而且在正常情况下,99. 5%以上的干细胞是处于G0静止期。造血干细胞具有不断地进行自我更新,维持在体内一定的数量和保持自己的特性,同时又具有向骨髓红系、粒系和巨核系祖细胞分化的能力。一个造血干细胞进行分裂后产生的两个子细胞,只有一个立即分化为早期祖细胞,另一个仍保持干细胞的全部特征不变。这种不对称性分裂,不论进行多少次,始终可以维持干细胞的数量不变,故能维持正常机体的长期、恒定造血。
造血干细胞的分化,受细胞与骨髓微环境、细胞表面、药物受体和环化酶系统以及体液等多种因素的控制和调节。造血干细胞的内在基因控制也起一定的作用。
红系祖细胞阶段
在红系祖细胞(progenitor cell)阶段,细胞是处于造血干细胞与红系前体细胞之间的细胞群。由红系祖细胞向红系前体细胞分化的阶段,是调节红细胞生成自体稳定机制中的一个关键过程。造血干细胞一旦变为早期祖细胞,立即出现对称性有丝分裂,其自我更新和自我维持的能力立即下降。晚期祖细胞则全部进行对称性有丝分裂,完全丧失了自我更新的能力。
造血干细胞在骨髓造血微环境的影响下分化为红系祖细胞。造血微环境包括微血管系统、神经系统和造血间质等部分。通过体液因子、细胞因子对造血干细胞的分化起特殊的作用和影响。红系祖细胞向红系前体细胞的分化是随机的系限过程。限制祖细胞只向单一红(红系)细胞发育。这种限制可能是由于细胞表面有系特异性生长因子受体的表达,如红细胞生成素(erythropoietin,EPO)受体等,此外也可能是由于某些因素与骨髓微环境相互作用的结果。骨髓微环境对造血干细胞的定向分化起着决定性的作用。如造血组织中血流量增加,组织氧分压增高,基质中的黏多糖倾向性变为中性,均有利于红系细胞的分化;否则,当组织氧分压降低、基质中的黏多糖倾向变为酸性时均不利于向红系细胞发育。
由于红系祖细胞可以在EPO的作用下向红系前体细胞的方向分化、增殖,最后成为成熟的红细胞,这类细胞也被称为EPO反应细胞(erythropoietin reaction cell,ERC)或EPO敏感细胞(erythropoietin sensitive cell,ESC)。这类细胞无自我维持能力,故不能称为干细胞。ERC或ESC细胞在高浓度的EPO条件下,当培养延续到14~16天,培养体系中会骤然生成由30 000~40 000个红系细胞组成的红系集落,称为红系爆式形成单位(burst forming uniterythroid,BFU-E),是ESC群体中较早期分化的细胞。BFU-E是更接近造血干细胞的红系祖细胞,可分化为红系集落形成单位(colony forming unit-erythroid,CFU-E),其比重沉降率较CFU-E缓慢。DNA合成期的比例亦较少,仅为0~25%。在形态学上较CFU-E更不成熟,呈轻度卵圆形,核染色体细,具有多个大的核仁,胞质呈碱性,偶有伪足。
BFU-E的数量为5~10/1×105有核细胞,与CFU-E不同的是BFU-E可见于周围血中,量极少,仅占0. 02%~0. 05%。
ERC或ESC在加入EPO的体外半固体培养环境中培养5~8天,可生成由8~65个红系细胞组成的细胞团,称为CFU-E。可以由G-6-PD同工酶分析确定。其形态学表现为核染色质细致,细胞核大,有大核仁,有清楚的核周带及少量的嗜碱性胞质。人类骨髓中的CFU-E数因不同的分离方法和培养条件而异,约为(50~400)×105/L有核细胞。大部分CFU-E是处于活跃的DNA合成期(S期),多数体外实验证实CFU-E细胞表面带有较密的EPO受体,且依赖EPO存活。
BFU-E和CFU-E是红系祖细胞群中两类性质不同的细胞亚群,它们的区别包括一般特性、对细胞因子的反应以及抗原和受体的表达。
红系祖细胞的一般特性及表面抗原/受体
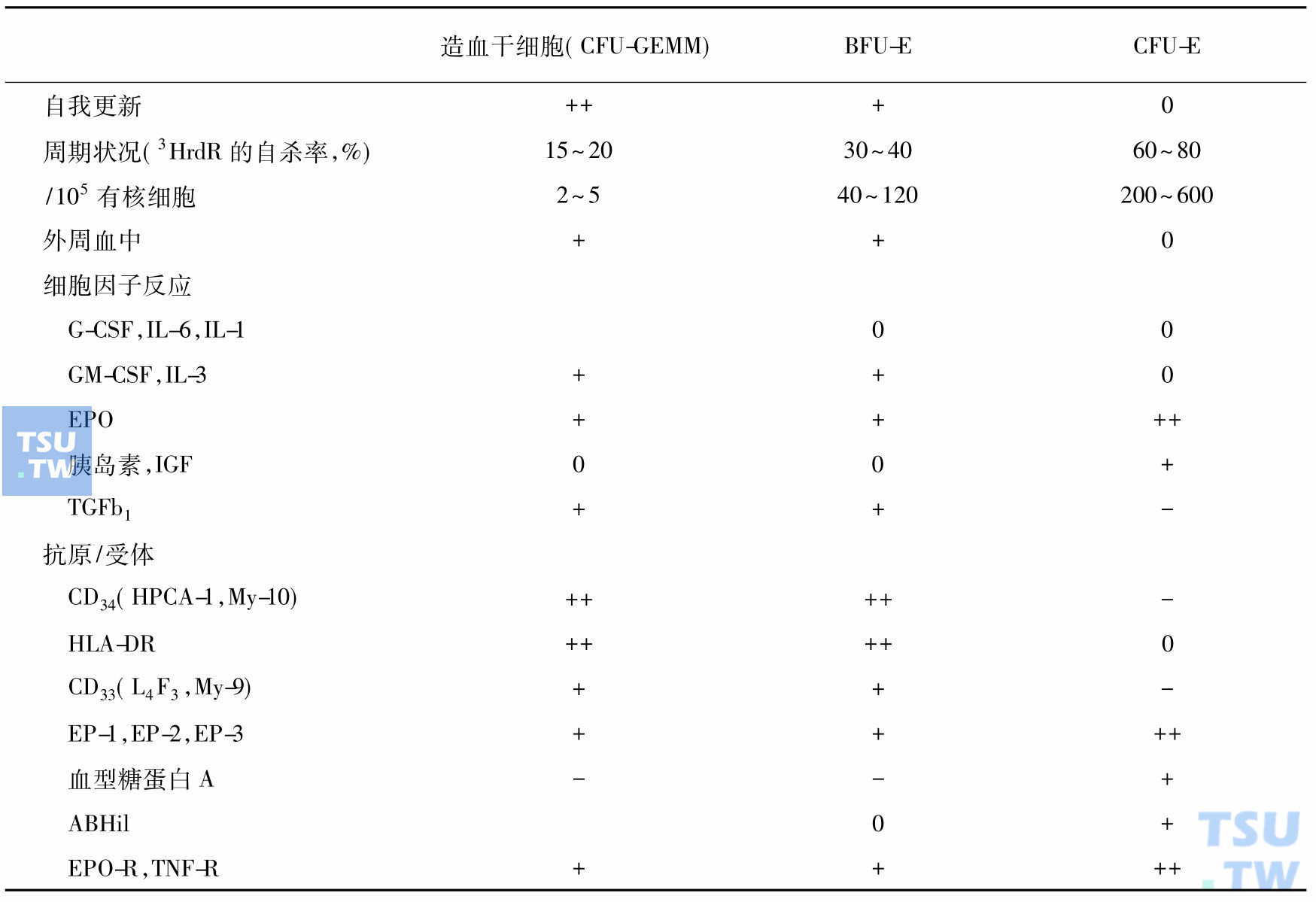
注:-表示缺如;+表示少量;++表示多量
主要影响BFU-E阶段的细胞因子是IL-3和GM-CSF。IL-3可以影响BFU-E的整个增殖期,其他如EPO、G-CSF等。在培养液中如无IL-3,BFU-E即不能生存。6天后,80%的BFU-E可消失。实验证实,早期阶段的BFU-E的增殖和分化均不依赖EPO,BFU-E在无EPO的环境下仍可存活48~72小时。晚期仅20%的BFU-E有低密度的EPO受体表达。对BFU-E负性影响的有α和β干扰素。后者可防止BFU-E进入细胞S期。
BFU-E进入CFU-E期后开始表达,可识别红系细胞的特征,这些具有特征的蛋白包括唾液酸糖蛋白、血型糖蛋白A(glycophorin A)和Rh抗原、血型抗原及ABHil型等。在CFU-E细胞上还存在大量的EPO受体。在缺乏EPO的培养液中数小时,80%的CFU-E细胞即不存在。EPO受体在CFU-E及原始红细胞上表达最多,以后随红细胞的成熟逐渐减少,到晚幼红细胞已消失。
转铁蛋白受体(TfR),也是在CFU-E和红系前体细胞时表达最高,到网织红细胞时最低。TfR是由双硫键连接的双链跨膜糖蛋白,分子量为180kD。每个受体可结合1个或2个分子的转铁蛋白。TfR是控制细胞摄取铁的重要因素,当红系细胞出现血红蛋白合成后,TfR的表达明显增多,随着细胞的成熟TfR逐渐减少。BFU-E和CFU-E上的TfR量较少,原始红细胞上的TfR最多,每个细胞可表达800 000个TfR,至网织红细胞,TfR的表达减少为100 000个/细胞,成熟红细胞则无TfR表达。
ERC(或ESC)是非均一的放大过渡细胞群体,随其放大而成熟。其数量控制着细胞进入CFU-E的速率。EPO在ERC晚期阶段可促成二级分化,生成下一级放大的过渡群体——红系细胞。其终末产物——成熟红细胞的数量通过组织氧分压控制着EPO的生成水平。当EPO缺乏时,ERC的G1期延长,红细胞的生成处于持续低水平。在贫血时EPO增加,ERC的G1期明显缩短,促进细胞进入S期,ERC池扩大,以适应加速红细胞造血的需要。
红系前体细胞阶段
红系前体细胞(precursor cell)阶段与BFU-E及CFU-E不同的是可以用形态学标准区分。包括原始红细胞、早幼红细胞、中幼红细胞、晚幼红细胞及网织红细胞阶段而达到成熟红细胞。细胞成熟的过程是血红蛋白增加和细胞核活性衰减的过程。随着细胞的成熟,有核红细胞中的血红蛋白含量不断增加,RNA的含量不断减少。在中幼红细胞后期,细胞中的血红蛋白含量≥13. 5pg。红细胞内血红蛋白的增高促使核失去活性,不再合成DNA或RNA。实验证实这是由于血红蛋白通过核膜间孔进入核内,作用于核组蛋白(nucleohistones),导致染色体失活而促进核凝缩。晚幼红细胞已失去继续分裂的能力,以后细胞核浓缩并逸出,被单核巨噬细胞吞噬,或在脾脏内碎裂、溶解,成为无细胞核的网织红细胞。在成熟红细胞阶段不再合成血红蛋白。
根据细胞内血红蛋白浓度的增高会促使细胞核失去活性的理论,红细胞成熟过程分裂的次数、细胞最终的大小与血红蛋白合成的快慢有一定的关系。如缺铁时的小红细胞是因为血红蛋白的合成速度慢,需要较长的时间才能达到需要的血红蛋白浓度,故在细胞核停止活动(或聚缩)之前分裂的次数多,造成细胞体积小。相反,在大细胞时,过早地使细胞核变性,分裂的次数也少些。
如果红系前体细胞由于某种原因在从骨髓释放前或以后短期死亡,称为无效造血。正常人的红细胞无效造血只占极少部分(<10%),而在某些病理情况(如巨幼细胞贫血、珠蛋白生成障碍性贫血及铁粒幼细胞贫血)时,无效造血会明显增加。红细胞无效造血的原因可能是:①红系干、祖细胞本身的缺陷,使生成的红细胞质和量异常;②幼稚红细胞的分裂周期延长,导致骨髓过度增生、成熟障碍,致使幼稚红细胞在骨髓内破坏;③有缺陷的红细胞生成及释放入血后不久即遭破坏;④骨髓幼稚细胞内造血物质(卟啉、铁)转输和代谢加速,出现“血红素无效生成”也可出现无效应红细胞生成。
决定红细胞外形及可变性的膜蛋白,如收缩蛋白、血型糖蛋白、带3蛋白、4. 1蛋白和锚蛋白均出现于CFU-E期。带3蛋白、4. 1蛋白及血型糖蛋白,特别是血型糖蛋白A,大量出现于CFU-E的晚期阶段或原始红细胞阶段。
随着细胞的成熟,红系细胞的直径逐渐缩短,细胞体积也逐渐缩小。这是因为细胞内一些用以合成血红蛋白、基质蛋白及各种酶的细胞器(如线粒体、高尔基器、聚核糖体)逐渐减少,细胞器也逐渐退化消失。
基因活性在红细胞成熟过程中是由珠蛋白的表达所支配。珠蛋白在原始红细胞中仅占蛋白质的0. 1%,到网织红细胞时达95%。对珠蛋白的研究及了解,在红细胞分子系列中是了解得最好的。已知成年人珠蛋白的合成主要是HbA(α2β2),少量的HbA2 (α2δ2)及HbF(α2γ2)。目前对Hb合成的数个酶基团已经克隆,如δ-氨基-γ-酮戊酸(δ-aminolevulinic acid,ALA)、胆色素原(prophobilinogen,PBG)脱氧酶及血红素(heme)合成酶等。
应用方向性核素标记细胞增殖周期DNA合成能力作指标可以推算红系细胞的增殖时间,原始红细胞的增殖时间约为20小时,早幼红细胞约为16小时,中幼红细胞为25~30小时,晚幼红细胞不具备合成DNA的能力,属非增殖性细胞。故正常红系前体细胞由骨髓生成,经过增殖、分化直到新生网织红细胞从骨髓中逸出约需3~5天。在贫血应激时,采用跳跃式分裂,此段时间仅可为2天。网织红细胞以后又在脾内停留1~2天,继续成熟且改变膜脂质成分后再进入血循环。
红细胞的脱核与释放
晚红细胞通过增加本身的波状运动,再经过几次收缩,把核挤到胞质的一极而后脱出。
网织红细胞通过骨髓-血液屏障亦是一个复杂的过程。骨髓血液屏障由一层薄的内皮和一层不连续的外膜组成。在骨髓的血窦间隙处尚有大量巨噬细胞分布(窦周巨噬细胞)它能吞噬脱出的红细胞核,筛过和移去缺陷细胞(约占2%)。
红细胞的释放是通过骨髓的窦壁、内皮细胞联合处的胞质而入血的。当红细胞进入血窦时,易变形的胞质先进入,把细胞核留在血窦处,红细胞进入血窦后,内皮细胞即收缩而使血窦孔闭合。
网织红细胞的成熟程度,可根据所含网状物量的多少分为Ⅰ~Ⅳ级。正常时仅Ⅲ及Ⅳ级网织红细胞见于外周血中。
