
趋化因子(chemokine)是指能使白细胞向炎症部位迁移的细胞因子,对调节白细胞的功能和运动方向起重要作用,主要与炎症、感染和免疫相关,近年来的研究进展表明,趋化因子影响造血干/祖细胞的迁移、归巢、动员、增殖和生存,参与造血调节。
趋化因子的结构特点:N端具有由至少三个β折叠结构和C端的α螺旋以及保守的四个半胱氨酸(C)形成的结构域:以N端的两个半胱氨酸结构为准,CXC族的趋化因子两个半胱氨酸间有另一任意氨基酸隔开;CC族的两个半胱氨酸直接相连;CX3C族的两个半胱氨酸间有三个氨基酸隔开,其C端有很长的附加结构,包括musin样结构;C族趋化因子很特殊,仅剩一个半胱氨酸,仅一个成员淋巴细胞趋化因子(lymphotactin)。
趋化因子是分子量8~12kD的蛋白。其原型是IL-8。至今已发现五十多种趋化因子,但是仅发现20种趋化因子受体。亦即趋化因子与其受体的结合不够特异,很少趋化因子受体呈特异性结合,如原来认为SDF-1α/CXCL12仅与CXCR4结合,后来发现也能与CXCR7/RDC1结合。趋化因子受体结构有别于其他细胞因子的受体结构,属七次跨膜型的G蛋白耦合受体(GPCR)。趋化因子基因的染色体定位如下,CXC族:14q12~21(SDF-1α例外在10号染色体);CC族:17q11. 2~12;CX3C族:16;C族:1q23。趋化因子基因染色体定位的聚集提示这些基因在进化上的同源性。根据趋化因子的结构和基因的染色体定位将受体分四类:CXCR:1~7;CCR:1~10;CX3CR:1和CCX-CKR;CR:1。
趋化因子的氨基酸序列类似,所以它们的二级和三级结构也相似。晶体分析表明:所有的趋化因子都是多聚体,除PF4是四聚体外均为二聚体,但是生理浓度下往往呈单体。由于趋化因子都是与细胞表面的受体或结缔组织中的氨基糖蛋白结合,二聚体化可能有优越性。
除了CCR6、CCR8、CCR9和CXCR5、CXCR6、CXCR7仅与一种趋化因子结合,其他趋化因子受体都与两种以上的趋化因子结合。许多趋化因子有一种以上的受体。为什么趋化因子多而受体少,同一趋化因子可结合多种受体。Weber等根据他们的实验结果作如下解释:激活不同的受体产生不同的功能。白细胞从血流迁移至炎症部位要经过四步:沿着血管壁滚动;停止滚动;播散;穿越内皮细胞进入炎症组织。Weber等发现在此过程中趋化因子激活两个不同的受体:CCR1和CCR5,CCR1在血流变缓时使白细胞停止滚动,CCR5则使白细胞沿着血管壁播散,两个受体均可使白细胞穿越内皮细胞进入炎症组织。趋化因子受体在人类疾病中起重要作用,从HIV感染、心血管疾病到自身免疫病都起十分重要的作用。若用抑制趋化因子的策略治疗这些疾病需针对大量受体。如果Weber等的假设成立就能减少许多治疗靶点。
趋化因子作用的复杂性与趋化因子-趋化因子受体相互作用的复杂性有关。趋化因子在炎症部位形成浓度梯度,低浓度处呈单体,高浓度处呈多聚体;趋化因子受体活化受细胞内状态和细胞聚集状态的影响,也能形成多聚体。趋化因子单体足以结合活化趋化因子受体,多聚体趋化因子对聚集的受体作用更强,表现在能募集更多活性更强的白细胞。SDF-1/CXCL12介导了众多功能是近年来的研究热点。
SDF-1/CXCL12功能的介导
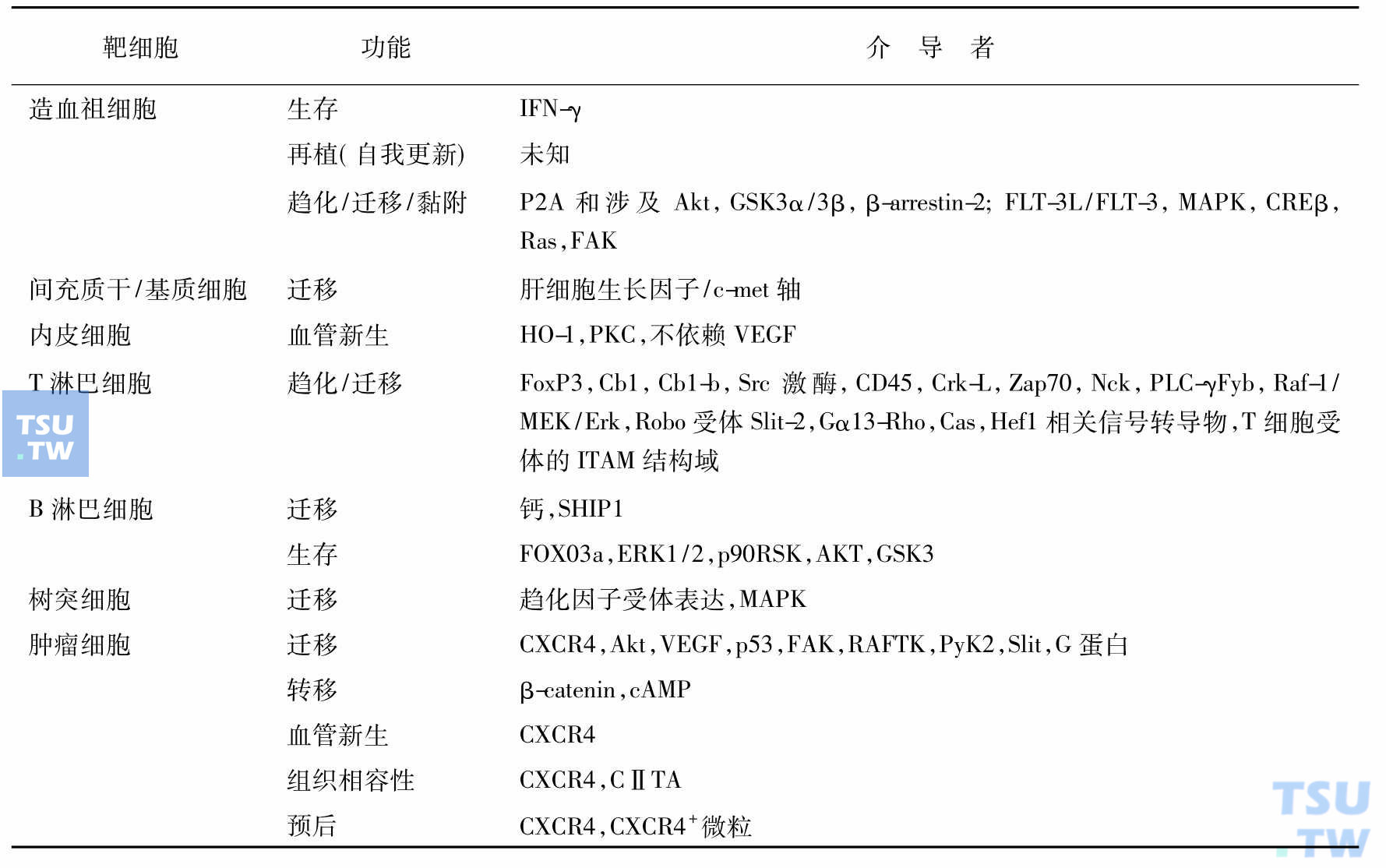
有人根据基因表达和受体种类,将趋化因子分为炎症趋化因子(inflammatory chemokine)和归巢(或稳态)趋化因子(homing/homeostasis chemokine)两大类。炎症趋化因子在大部分组织不同类型的细胞和迁移来的白细胞在炎症细胞因子、细菌毒素和其他病理因素作用下产生,除中性粒细胞激活的多肽2(NAP2)之外,CXCR1、CXCR2、CXCR3、CCR1、CCR2、CCR3和CCR5的所有配体均属此类。归巢趋化因子则由淋巴组织呈构成性表达,它们与淋巴细胞的归巢和成熟有关。趋化因子/趋化因子受体与造血调节的关系,还有许多有待研究阐明之处,如趋化因子样受体(chemokine-like receptor,CMKLRs)和非趋化因子与趋化因子受体的结合使问题复杂化,有待深入研究。
