
近年来,一类核糖核酸小分子(即microRNA 或miRNA)成为全球生命科学、生物医学、药物学等诸多领域关注的焦点。具有调节功能的非编码小分子RNA是指一类内源性长度为21~24nt的单链核酸小分子,通过与靶基因3'-UTR区的完全或不完全互补结合,从而引起mRNA的降解或翻译抑制发挥负性调节作用。小分子RNA大多在进化上具有序列保守性,不编码蛋白质,而是以RNA的形式,通过调节一些信号分子,如生长因子、转录因子、前程序性细胞死亡基因和抗程序性细胞死亡基因的表达,实现对细胞死亡、增殖、分化、发育和新陈代谢的调控。小分子RNA的发现不仅揭示了困扰科学家们多年的基因沉默现象的本质,还改变了人们对基因的传统认识。miRNA的发现是近年来生命科学研究的一个重大突破,在2002年和2003年连续两年被《Science》杂志评为“年度十大科技进展”。
miRNA的生成主要包括两个过程。首先在细胞核内编码miRNA的基因由RNA聚合酶Ⅱ(RNA polⅡ)转录生成单顺反子或多顺反子初级转录物(Pri-miRNA)。Pri-miRNA随后进入微处理器Drosha(一种RNaseⅢ酶),在此被切割成60~70nt的一端5'磷酸,一端3'端两个悬垂结构的miRNA前体——pre-miRNA。pre-miRNA在Ran-GTP依赖的核质/细胞质转运蛋白Exportin5的作用下,从细胞核运送到细胞质中,然后被另外一种RNaseⅢ核酸内切酶复合物剪切成21~25个核苷酸长的miRNA:miRNA*双体。此复式结构在解旋酶作用下,一条链被降解(通常是miRNA*),而另一条链(通常是miRNA)进入核蛋白复合体形成RISC(RNA—induced silencing complex,RNA诱导的沉默复合体),发挥生物学作用。
根据与靶基因互补程度的不同,miRNA作用机制分为两种:靶mRNA切割和翻译抑制。miRNA同蛋白编码mRNA完全或几乎完全互补时,通过RNA介导的干扰途径,即miRNA相关的核糖核苷酸和miRNA诱导的沉默复合物(RNA induced silencing complex,miRISC)联合作用,降解靶mRNA。大多数动物体内,miRNA并不直接降解靶mRNA,而是与其3端非翻译区(3'-UTR)不完全互补,同样通过RISC或RNA干扰途径明显抑制转录后翻译水平的基因表达。miRNA运用此机制亦能降低相应靶基因的蛋白表达水平,不影响mRNA本身。
对于miRNA的最早研究是1993年Lee等用正向遗传学的方法在秀丽线虫中发现了第一个miRNA lin-4;lin-4有两种形式,一种是60nt长度,另一种是22nt的长度,均可形成茎环状的发夹结构,但22nt长度的可以通过与lin-14 mRNA 3'-UTR以不完全匹配的形式抑制lin-14基因的表达。2000年Reinhart等在秀丽线虫发现了另一个miRNA lin-7。lin-7的发现具有重要的意义,因为,不同于lin-4,lin-7在多种生物中表达,有较强的保守性,说明miRNA介导的基因调控具有比以往想象的更古老也更普遍的规律。随后,科学工作者采用多种方法寻找miRNA分子,迄今在人类组织中发现大约六百多条此类分子。而预测人体内约有一千条这样的小分子,它们调控约1/3人类基因的翻译表达。
miRNA在正常造血中的作用
造血是多层次复杂调控过程,涉及造血干细胞自我更新和向不同形态及功能的血细胞分化、成熟。这些过程受严格的调控,这些调节机制的分子性质尚知之甚少,目前的研究成果多集中于转录因子的转录调控。随着miRNA的发现,人们越来越重视这一新的调控层次,miRNA介导的转录后调控既可作用于转录因子直接参与调控过程,又可作用于某些功能蛋白作为转录调控的补充。
miRNA与造血有关的最初证据来自于Chen等的研究,他们从小鼠骨髓克隆了大约100个miRNA,发现其中三个(miR-223、miR-142、miR-181)主要表达于造血组织。进一步鉴定发现,miR-181于胸腺高表达,在未分化细胞(Lyn-)低表达,在B淋巴细胞中表达上调。在造血祖细胞中异位表达miR-181,将这种细胞移植入致死照射的小鼠,会造成B系淋巴细胞比例的显著升高(较对照高53%)以及T细胞比例(Thy-1. 2+)的显著降低(88%)。随后的研究证实,不仅造血组织和非造血组织miRNA表达谱大不相同,在造血组织内部也存在差别。miRNA与造血干细胞向各系定向分化有关,并可能对分化起调控作用。
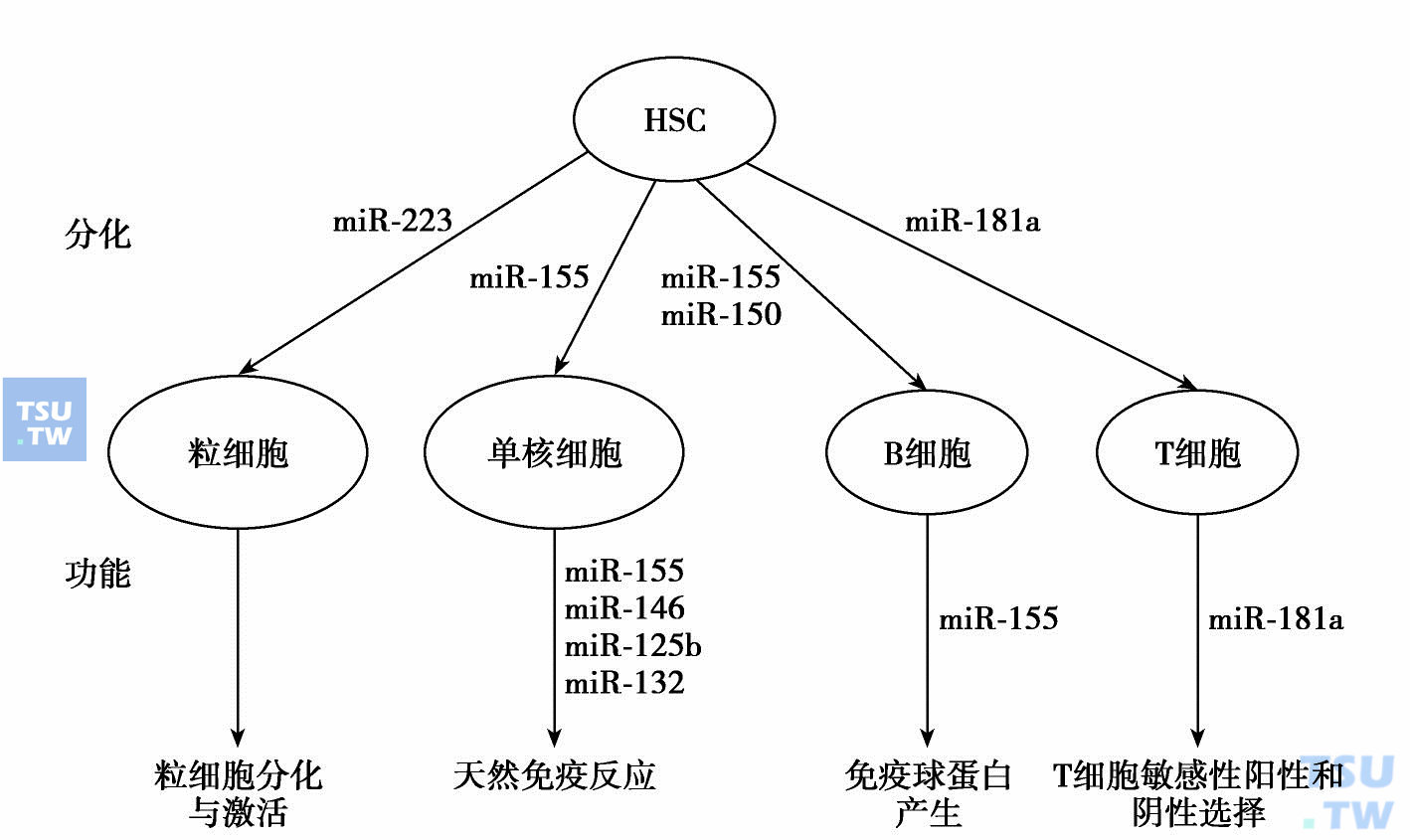
miRNAs参与造血分化和功能调节
淋巴细胞发育
研究发现在T细胞分化过程中,miR-181于胸腺高表达,并在胸腺CD4+CD8+双阳性细胞中表达上调。在这一发育阶段中miR-181a的上调与bcl-2、CD69和TCRα的表达呈负相关。进一步的研究表明,miR-181家族成员在胸腺发育中靶向bcl-2、CD69和TCRα调节其表达水平。由于CD69信号的破坏会影响淋巴细胞从胸腺的移出,异位表达的miR-181可能越过CD4+CD8+双阳性阶段,引起低表达CD69的细胞在胸腺中滞留,从而导致外周T细胞的减少。
miR-150选择性表达于成熟静息B细胞和T细胞,而在B、T祖细胞中不表达。过表达miR-150的转基因小鼠B细胞明显减少,表明miR-150通过靶向转录因子c-MYB调控B细胞分化。
miR-17~92基因簇也对B细胞发育至关重要。其在小鼠中的靶向缺失导致原凋亡蛋白Bim水平升高,抑制原B细胞向前B细胞的发育。这些证据表明miR-17~92基因簇在原B至前B细胞的发育转换中起特异性调节作用,通过靶向原凋亡蛋白Bim,促进这一阶段B细胞的存活。
miRNA与免疫系统
miRNA不仅在B、T淋巴细胞不同发育阶段具有不同的表达特征和调节功能,而且在调节免疫反应方面起重要作用(下表)。采用miR-155缺陷小鼠实验显示,miR-155调节生发中心功能、T细胞依赖抗体反应和细胞因子产生,对正常B、T细胞发挥功能至关重要。
参与免疫系统发育和功能调节的miRNA
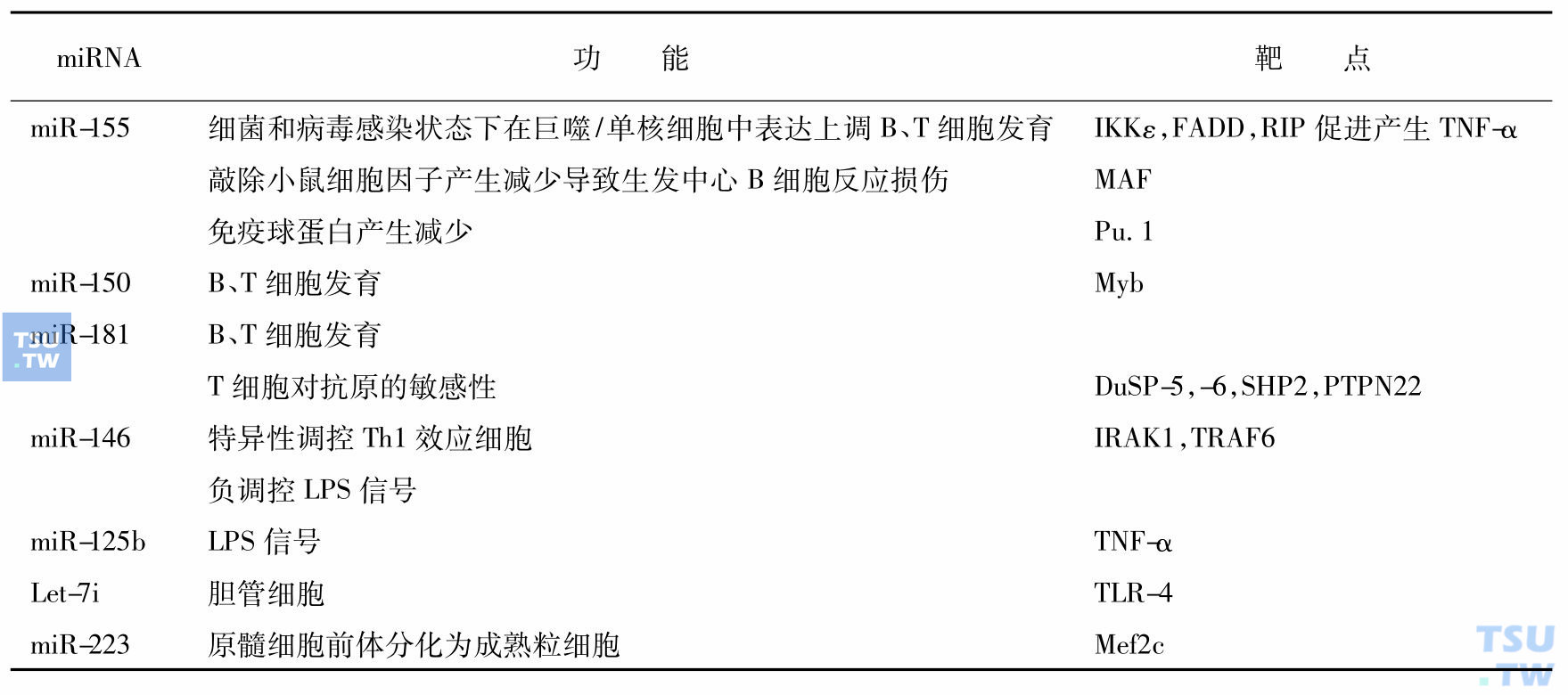
进一步的实验揭示了miRNA在精确免疫调节中的作用。miR-181a可在发育过程中调控T淋巴细胞受体的敏感性。成熟T细胞中miR-181a表达升高增强T细胞对肽抗原的敏感性,而抑制不成熟T淋巴细胞中的miR-181a会降低其敏感性并破坏阳性和阴性选择。这些作用部分源于miR-181a对多种磷酸化酶(SHP-1、SHP-2、DUSP5和DUSP6)的多靶点调控,引起高水平磷酸化中间产物的积累,从而降低TCR信号的反应阈值。
巨核细胞与红细胞发育
在CD34+造血祖细胞(HPCs)向巨核系定向分化的同时伴有20种miRNA的下调。其中miR-10a和miR-130a可下调mafB和HOXA-1,而这两个基因均在此分化过程中上调,表明miRNA能够阻断它们的表达。在红系,CD34+HPCs向红系分化过程中出现miR-221和miR-222下调。这两种miRNA在CD34+HPCs中的异位表达抑制红细胞生长和Kit蛋白表达。进一步的实验揭示,miR-221和miR-222的下调可解除对kit表达的阻断作用,促进早幼红细胞的扩增。对红系前体细胞进行miRNA表达的系统分析显示了分化晚期miR-150、miR-155、miR-221和miR-222的进行性下调,miR-16、miR-451的上调,以及miR-339 和miR-378的双相调节。
采用条件GATA-1细胞系,Dore等发现miR-144/451基因簇处于红细胞关键调节因子GATA-1的转录调控之下。采用反义吗啉代寡聚体干扰斑马鱼胚胎中该miR基因簇的表达,发现注射miR-451反义寡聚体的胚胎有正常的红系前体,但向成熟红细胞的发育被破坏。注射miR-144反义寡聚体不引起任何明显改变。
粒细胞和单核细胞发育
研究发现,miR-223在CD34+HPCs和髓系前体细胞中低表达,随着粒细胞分化,其表达稳步增加,而在单核系中表达受抑。与此一致,miR-223在急性早幼粒细胞白血病(APL)细胞系和患者标本用维A酸体外诱导粒系分化过程中表达上调。在寻找miR-223调节元件过程中,Fazi在miR-223前体-709碱基处发现了一个非保守的C/EBPa结合位点。采用APL细胞系进行的进一步实验表明C/EBPa与转录因子NFIA竞争结合这一位点。用维A酸处理APL细胞引起NFIA取代C/EBPa并导致miR-223的转录激活,miR-223的激活反过来抑制NFIA的转录。此外,在APL细胞中异位表达miR-223可以促进粒细胞分化。因此,从上述结果看,miR-223对粒系生成起正向调节作用。而其他研究组在小鼠和人的miR-223启动子位点的更远端发现了转录因子PU. 1和C/EBPb的结合位点,这两种转录因子与这一位点结合可激活miR-223的转录。并发现,APL细胞用维A酸诱导分化过程中抑制PU. 1可强烈抑制miR-223表达,因此认为在这一模型中,miR-223的表达主要由PU. 1驱动。但是,Johnnidis发现miR-223缺陷小鼠循环以及骨髓中性粒细胞的数量显著增加,这些粒细胞形态异常,粒系表面标志表达异常,并对刺激信号反应性增强,出现自发性炎性肺病。此外,他们还发现,促进髓系祖细胞分化的转录因子MEF2c是miR-223的靶点,剔除Mef2c基因后,miR-223缺陷小鼠表型恢复正常。因此,认为miR-223的作用是减少粒细胞产生并破坏其激活。总之,有关miR-223在粒系生成中的作用尚存争议。
在单核细胞发育中,PU. 1激活miR-424,在急性髓系白血病(AML)细胞系和CD34+HPCs中,激活的miR-424通过抑制转录因子NFIA反过来诱导单核/巨噬分化。
miRNA在不同的造血细胞分化阶段表达不同可能是由于miRNA转录生成pri-miRNA时受到特异性调控细胞分化的转录因子调节,也可能是miRNA在转录后调控某些系列特异基因的表达,参与造血分化决定。比较在造血分化过程中不同系列分化时转录因子及特异基因的变化,与相应的miRNA的变化相联系,再根据miRNA靶基因预测的结果和miRNA上游转录因子结合序列可能进一步揭示miRNA调控网络。
miRNAs与白血病
白血病是以异常血细胞增殖为特征的骨髓克隆性疾病。白血病患者中普遍存在非随机的染色体异常(如缺失、易位、倒位)导致的不可修复的基因功能缺失(如失去肿瘤抑制基因)或癌基因激活,上述因素一直被认为是导致白血病发生的关键。
慢性淋巴细胞白血病
在慢性淋巴细胞白血病(CLL),最为常见的染色体异常是13q14区的杂合或纯合或双染色体缺失,发生率超过50%,并与发病相关。miR15、16-1正好处于CLL基因缺失区13q14的30kb内。与健康供者正常成熟CD5+淋巴细胞相比,在大多数(68%)B淋巴细胞性白血病(B-CLL)患者的miR-15、16-1表达下调。在两例CLL患者白血病细胞中找到了引起等位基因缺失从而造成这两个miRNA基因簇表达沉默的相关突变。此外,在自然发生的迟发型性CLL小鼠模型中发现miR-16-1附近3' DNA的点突变导致miR-16-1表达降低。因此,不仅仅是染色体缺失,点突变也会引起CLL miR-15a/miR-16-1的表达降低。有趣的是,miR-15a/16-1的靶基因是癌基因bcl-2,原发性CLL患者中miR-15a/16-1的表达与bcl-2蛋白表达呈负相关。CLL恶性细胞大多过表达bcl-2,bcl-2本身具有抗凋亡作用,能够使肿瘤细胞逃避细胞凋亡。miR-15a和miR-16-1通过直接与bcl-2 mRNA的3'-UTR序列相互作用调控bcl-2蛋白的表达,以类似于抑癌基因的作用抑制B淋巴细胞的过度增殖,而它们的缺失则导致B细胞增殖能力大大增强,从而脱离正常的增殖凋亡平衡走向恶性转化的道路。体外实验证实miR-15a和miR-16-1在转录后水平抑制bcl-2的表达足以引起CLL来源细胞系的凋亡,因此,miR-15a和miR-16-1作为天然的bcl-2“抑制剂”有望应用于临床治疗CLL及其他bcl-2过表达引起的肿瘤。Calin等在一个家族性CLL并同时伴有乳腺癌的家系中鉴定出了miR-15a和miR-16-1基因的单核苷酸突变,该突变直接导致了miR-15a 和miR-16-1的表达下降,同时在75例CLL患者中的11个检出了同样的突变,因此,小RNA基因突变有望成为家族性CLL或其他癌症与CLL并发率的可能指标。
另外,两个在CLL发病中起关键作用的miRNA 是miR-29b和miR-181b。这些miRNA通过靶向调节TCL1发挥肿瘤抑制作用。TCL1是共激活AKT并参与调节细胞存活、增殖和死亡相关多条通路(如mTOR、NFκB、mdm2和cyclinD1通路)的癌基因。高水平的TCL1与ZAP-70以及非突变IgVH高表达相关,而后二者是侵袭性CLL的标志。转基因小鼠实验亦证实,TCL1能诱导IgV区重排,而IgV区重排正是CLL中侵袭性亚型的典型特征。
急性淋巴细胞白血病
到目前为止,急性淋巴细胞白血病(ALL)患者标本miRNA表达情况及功能的研究很少。Mi等发现了27个表达异常(6个上调,21个下调)并且在AML和ALL中有表达差异的miRNA。其中,miR-128b在ALL中的表达高于AML,并大大高于正常CD19+细胞。
急性髓系白血病
对240个具有中危至高危细胞遗传学异常的急性髓系白血病(AML)标本的miRNA表达进行分析,发现miRNA表达谱与FLT3-ITD相关,表明这类患者细胞遗传学异常可导致miRNA的表达。其中,miR-191和miR-199a的过表达与AML患者总体生存时间和无病生存时间呈负相关。
在对携带NPM1和FLT3-ITD突变的AML患者miRNA的功能研究中发现,这两类突变通常出现于正常核型的AML患者中,约占AML患者总数的30%~40%。其中NPM1突变患者miRNA表达谱与NPM1非突变者有明显不同,主要表现为前者有miR-10a、miR-10b以及let-7和miR-29家族成员的表达上调。表达下调的miRNAs中miR-204靶向调节HOXA10和MEIS1,表明NPM1突变的AML患者HOX表达上调可能部分源于调控HOX的miRNA的缺失。FLT3-ITD+标本以miR-155上调为特征。进一步的实验证实,miR-155的上调不依赖于FLT3信号。给C56BL6小鼠移植过表达miR-155的小鼠HPCs,C57BL6小鼠骨髓出现伴有红系/巨核系减少的髓系增殖性疾病。这一结果与CD34+祖细胞向红系和巨核系分化过程中出现miR-155下调,以及在HPCs中过表达miR-155会阻断这两系体外分化这一现象相吻合。上述资料表明,miR-155阻断红系/巨核系分化,并与FLT3-ITD发病相关。由于miR-155的表达独立于FLT3信号,在治疗这一类型AML时应考虑用miR-155反义寡核苷酸阻断miR-155的同时联合使用FLT3-ITD抑制剂治疗。
在对APL细胞用维A酸(RA)诱导粒系分化过程中发现有miR-223特异性上调,而miR-181或miR-142表达水平未受影响。稳定过表达miR-223会促进RA反应性髓系白血病细胞系NB4向粒系分化,而干扰miR-223则抑制NB4对RA诱导分化作用的反应性,表明miR-223表达水平与髓系前体的分化密切相关。miR-223转录后调控对粒细胞分化以及APL白血病细胞对全反式维A酸的临床反应至关重要,为APL的诊断和治疗提供了新的靶点。
慢性粒细胞白血病
有研究表明,慢性粒细胞白血病(CML)细胞系中BCR-ABL和c-MYC均可以反式激活miR-17~92基因簇的上调。BCR-ABLMYC通路对pri-miR-17~92表达的诱导作用主要出现在慢粒慢性期的早期,而非急变期。
淋巴瘤
miR-155在很多人类肿瘤包括弥漫性大B细胞淋巴瘤(DLBCL)、原发性纵隔B淋巴瘤(PMBL)、Hodgkin淋巴瘤以及儿童Burkitt淋巴瘤(BL)中过表达,并发挥癌基因作用。在靶向B细胞过表达miR-155的转基因小鼠模型中,首先出现多克隆的前白血病前B细胞增殖,随后发生B细胞恶性肿瘤。miR-155在具有激活B细胞亚型中出现,而在幼稚B细胞亚型中不出现,而前者比后者的侵袭性强,因此miR-155可能成为DLBCL的临床诊断指标;在BL中miR-155的表达比正常人高出100倍,提示miR-155可能在BL的发生和发展过程中起作用,其机制可能是通过间接抑制某些抑癌基因的表达而实现。
13q31~32区的异常扩增在B细胞淋巴瘤患者中较为常见。而miR-17~92基因簇(包括miR-17-5 p、miR-17-3p、miR-18a、miR-19a、miR-20a、miR-19b-1 和miR-92-1基因)恰恰定位于这一区域,导致65% 的B细胞淋巴瘤患者miR-17~92表达水平明显增加。在MYC转基因小鼠的HPCs中过表达miR-17~92基因簇可以加速肿瘤形成。在淋巴细胞中过表达miR-17~92会使小鼠发生淋巴增殖性疾病、自身免疫以及成熟前死亡。miR-17~92基因簇通过下调PTEN和调节B细胞凋亡的原凋亡蛋白Bim促进淋巴细胞增殖和存活。深入研究发现,MYC可以结合并激活位于染色体13q31区域的miR-17~92的表达。此外,这一基因簇中两种miRNAs:miR-17-5p 和miR-20a可下调E2F1转录因子,而E2F1可促进细胞周期进展,是MYC的直接下游靶点。因此,MYC通过同时激活E2F1并通过基于miRNA的抑制机制限制其翻译,从而实现了对细胞增殖的紧密调控。
侵袭性淋巴细胞瘤白血病中常有t(8;17)染色体的异位,miR-142基因正好位于这个区域。异位使miR-142基因与c-Myc基因融合,导致c-Myc在miR-142基因启动子控制下高表达,促使低度恶性的慢性淋巴瘤演变为高度恶性的B细胞性白血病。
结语
最初在对秀丽线虫进行遗传筛选中发现的miRNA正引起越来越多的关注,成为人类遗传领域的新星。已有明确证据表明miRNA参与了造血细胞的发育和功能调控。miRNA精密调控造血细胞分化并调节细胞对刺激的反应性。其作用可能源于其对信号通路相关蛋白的多靶点协调抑制。在白血病和淋巴瘤,已发现广泛的miRNA调节异常。但是,完全阐明miRNA在造血和肿瘤中的作用还有大量工作要做。了解miRNA与转录体的相互作用,是阐明miRNA对基因表达的调控方式以及设计基于miRNA的治疗策略中重要的一环。
