
免疫应答过程中T细胞的激活需要双信号。第一信号来自TCR识别MHC/抗原肽复合物,传递抗原特异性识别信号;第二信号由APC的共刺激分子提供,为非特异性协同刺激信号。大量实验证实,如果缺乏共刺激分子提供的协同刺激信号,T细胞便不能激活,而处于无能(anergy)状态,甚至发生凋亡。
T细胞通过TCR识别MHC/抗原肽后,出现一系列和激活有关的事件。
信号跨膜传递和转导通路的启动
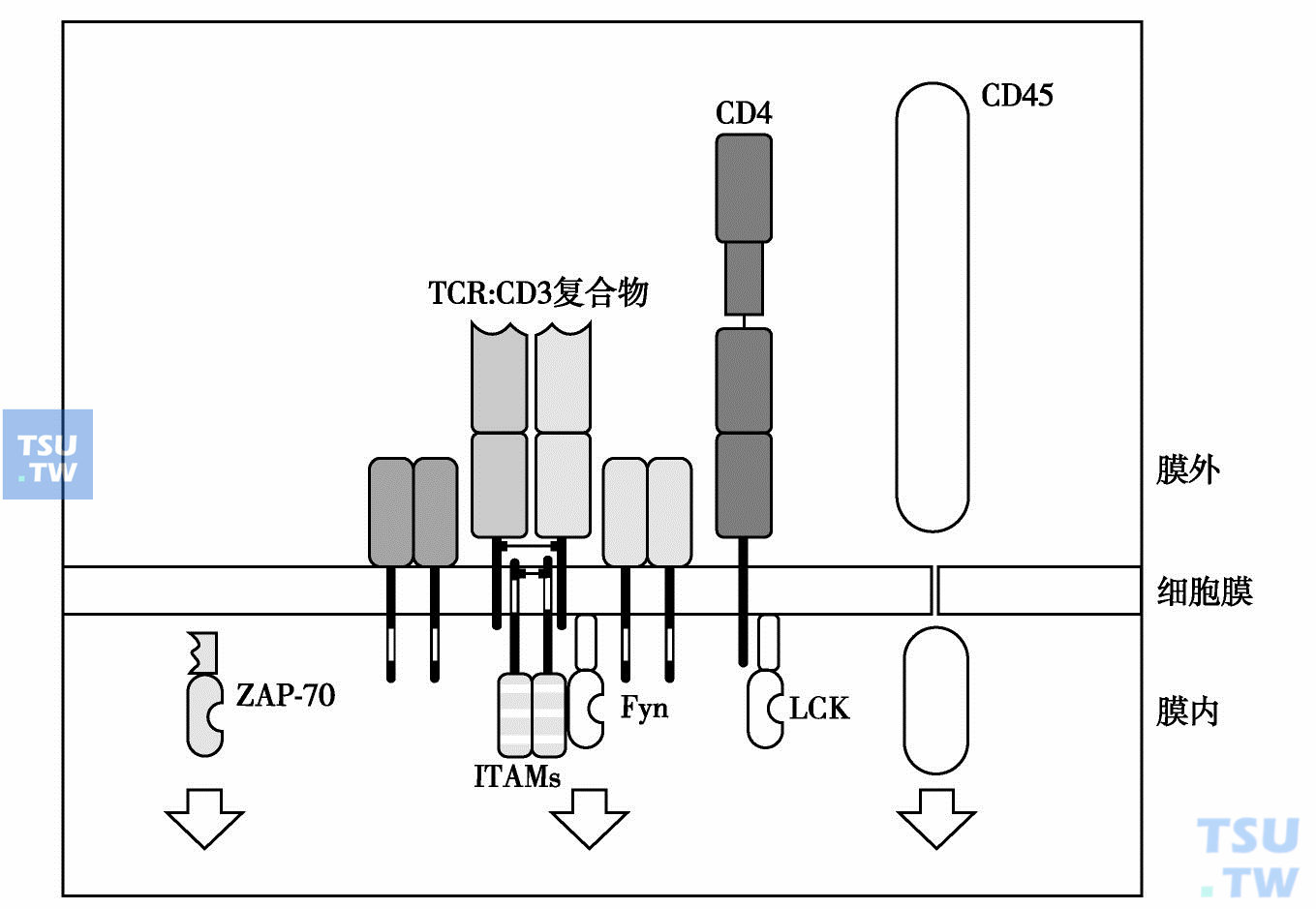
TCR和MHC/抗原肽的结合,引起细胞膜上相关分子的多聚作用(下图)。参与T细胞激活的多种跨膜分子如TCR-CD3、辅助受体CD4或CD8、CD45等相互靠拢成簇。细胞内分别和上述受体分子关联的蛋白酪氨酸激酶(PTKSrC)P59fyn分子(主要与CD3ζ链和CD3 ε链相连)、P56LCK(与CD4或CD8相连),以及蛋白酪氨酸磷酸酶(PTPase)CD45相互靠近。PTPase解除PTKSrC分子的抑制作用。胞内PTKSrC分子(fyn-LCK)相互磷酸化而被激活。
激活的PTKSrC使CD3分子(主要是CD3ζ链)胞内段上的免疫受体酪氨酸激活基序(ITAM)中的酪氨酸发生磷酸化;磷酸化的ITAM借助和SH2 (SrC同源区2)结合,招募PTK SYK家族中的重要成员ZAP-70;然后,已活化的SrC使得招募至CD3ζ链附近的ZAP-70分子发生磷酸化而激活,从而引起跨膜的T细胞活化连接蛋白LAT胞内段上多个酪氨酸残基发生磷酸化。磷酸化的LAT把各种带有SH2结构域的信号蛋白招募至TCR CD3附近,其中包括胞膜内侧的磷脂酶C(PLCγ)和转接蛋白Grb2。后两者启动(或参与启动)两条不同的信号转导途径。
参与T细胞激活的多种跨膜和膜内分子
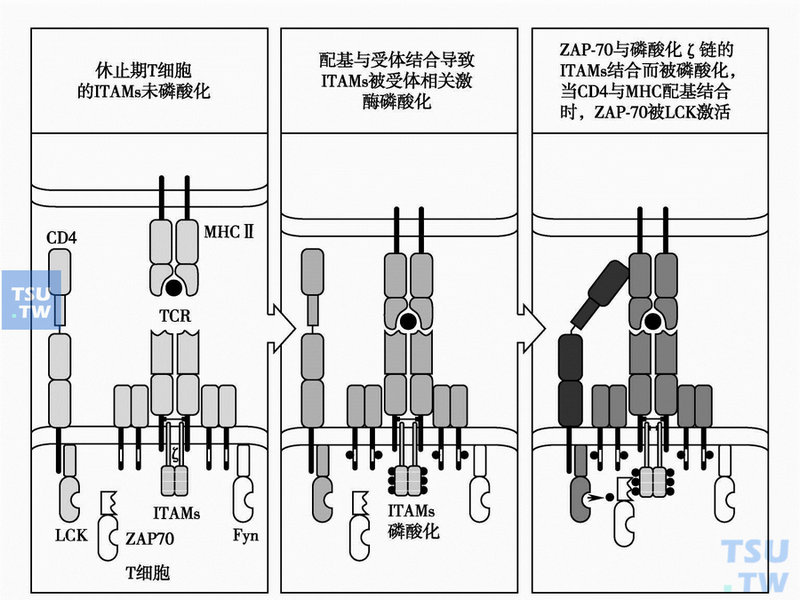
TCR与MHCⅡ类-抗原肽结合和辅助受体多聚启动T细胞内信号转导
从上可见,在T细胞信号转导起始阶段发挥作用的成分主要有四种:LCK/Fyn(属PTKSrC)、CD3ζ链、ZAP-70(属PTKsyk)和连接蛋白LAT。CD45分子(PTPase)以特有的方式解除SrC的抑制状态,直接参与T细胞激活信号通路的启动。胞内SrC成功地发生磷酸化表明PTK开始激活,胞外受体对抗原的识别有效。
抗原激活信号胞内转导的主要途径
磷脂酰肌醇途径(PI途径)
首先由激活的ZAP-70和PTKSrC使膜结合的磷脂酶C(PLC)分子γ链上的酪氨酸残基发生磷酸化。磷酸化的PLCγ发挥酶活性,使底物二磷酸磷脂酰肌醇(PIP2)水解成三磷酸肌醇(IP3)和甘油二酯(DAG)。IP3可迅速从膜内侧向胞质中扩散,一方面打开细胞膜上钙通道使钙离子进入细胞内,同时开启细胞内钙池增加钙离子的释放,协同提高胞内游离钙的浓度,激活钙调素(calmodulin)的Ca2+结合蛋白,调节其他酶类的活性,最终导致钙调磷酸酶激活。另一方面,和膜内侧相结合的DAG则直接激活蛋白激酶C(PKC)。钙调磷酸酶和PKC继之分别活化两种重要的转录因子NF-AT和NF-κB。实际上,这一条信号转导的下游通路中再一分为二,形成钙调磷酸酶参与的途径和PKC介导的途径,这就放大了传入的抗原识别信号,保证其转导的有效性。
MAP激酶相关途径
激活的ZAP-70使连接蛋白LAT和SH2结构域的白细胞磷酸化蛋白(SLP76)发生磷酸化,后者募集和激活其他的转接蛋白如生长因子受体结合蛋白2(Grb2)和鸟苷酸置换因子Sos,启动T细胞活化信号转导的第二条途径。在原癌基因产物Ras蛋白参与下,通过MAP激酶中的ErK和JnK激活Jun和Fos,构成转录因子AP-1。
T细胞激活的第二信号
前已提到,未致敏的T细胞通过TCR识别MHC-抗原肽获得抗原识别第一信号后,必须有共刺激分子提供的协同刺激信号(costimulatory signal)——第二信号,T细胞才能被激活(下图)。没有第二信号,许多基因,尤其是编码IL-2的基因不发生转录激活,没有IL-2的作用,获得了抗原识别信号的T细胞不能进入增殖分化阶段,呈现无能状态,还可发生凋亡。
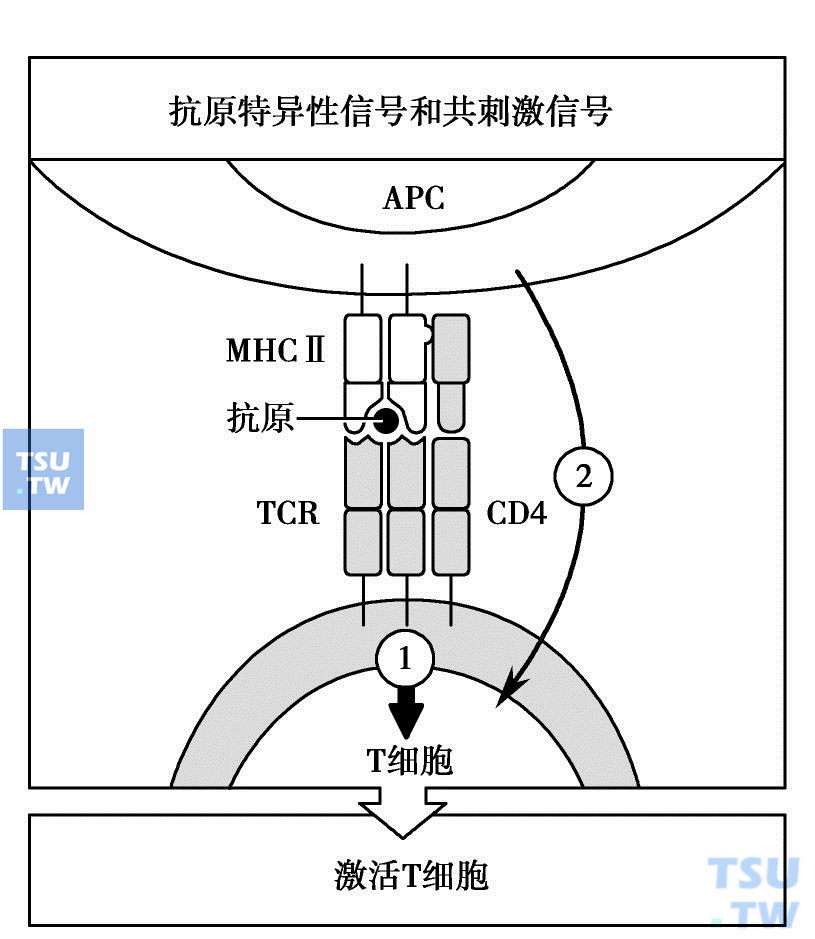
天然T细胞激活需要两个独立的信号
第二信号可有多方面来源,主要来自专职APC表面的B7分子(配体)和T细胞上的CD28分子(受体)的结合。第二信号不具有抗原特异性。专职APC中的DC可组成性地提供第一信号和第二信号,因此可以有效地激活未致敏的T细胞。而未激活的单核巨噬细胞和B细胞都不表达共刺激分子,它们只能提供第一信号,不能激活T细胞,相反可能引起T细胞无能或耐受。但一旦单核-巨噬细胞和B细胞被激活,它们都可以诱导性表达高水平的MHC和共刺激分子,有效地递呈抗原和使T细胞激活。如果T细胞识别其他细胞(如非专职APC)提供的MHC/抗原肽,往往因缺乏第二信号而进入无能状态,以后即使再有来自专职APC提供的第二信号,亦难以诱导IL-2基因表达,T细胞不再激活。
其实,这并非完全是坏事,因为这种情况下T细胞识别的往往是自身抗原。虽然前已提到能与自身抗原起反应的T细胞克隆已在胸腺发育中通过阴性选择被清除,但由于某些自身抗原仅表达在特定组织细胞上而不表达于胸腺基质细胞,因而相应T细胞克隆仍可存在,它们离开胸腺后分化成熟。一旦被激活,有可能损伤自身组织。因此,使这类识别自身抗原的T细胞进入无能状态或发生凋亡,有利于自身耐受的维持。然而,表达MHCⅠ类分子/抗原肽的肿瘤细胞虽可被CTL识别,却不能使之激活。因为肿瘤细胞(靶细胞)大多缺乏提供协同刺激信号的能力,这是肿瘤免疫治疗面临的一个重要问题,即如何防止免疫耐受。目前看来,必须依靠专职APC,尤其是DC。因此,如何有效地使负载肿瘤抗原肽的DC用于肿瘤免疫治疗的研究,将对包括肿瘤免疫学在内的整个免疫学的发展具有十分重要的意义。
