物质运转
机体的生存依赖于细胞内外的物质交换,细胞这种功能主要由质膜行使,不论外界发生什么变化,细胞都能够保持内环境的恒定。如何维持细胞内环境的恒定,方式有多种多样:
简单扩散
也就是通常说的高浓度向低浓度扩散。扩散的速度与浓度成正比,细胞外浓度愈高扩散速度愈快。因为质膜是脂质双层结构,各种不同的溶质,进入膜内的速度不同。一般情况是极性弱的,脂溶性较强的分子容易通过,反之,极性愈强,脂溶性愈弱而水溶性强的难在膜内扩散。非脂溶性的物质,分子愈小愈容易通过,分子大的很难通过。物质的解离度也有影响,一般电解质比非电解质难通过,因为电解质解离时外面有水化层,体积加大,所以难以通过。一般来说,机体以这种方式进行运转很少。
促进扩散
促进扩散即是需要一个载体帮助运转。载体有特殊的结构,运输不同的物质有不同的载体,如缬氨霉素是钾离子的载体,特异运钾离子,对钠离子的运输很慢。现在发现许多抗生素都是离子的载体,如镰孢菌素、短杆菌肽可能都是离子载体,它插入细菌的膜使细菌的细胞内外离子失衡而死亡。
主动转运
细胞内外的物质浓度往往是逆差,要把细胞外低浓度的物质向细胞内高浓度的方向运输,是个需能反应。如钾离子,细胞内浓度高,需从细胞外输送;钠离子在细胞内浓度低,需从细胞内运出去,膜上有一种酶称钾-钠ATP酶(Na+-K+ ATPase),这个酶的底物是ATP,每分解一个ATP,可泵出3个Na+,输入2个K+,所以钾钠ATP酶又称钾钠泵,它的反应速度很快每分钟1000次,用它来维持细胞内钾高钠低的正常生理情况。除钾钠泵外还有Ca2+泵及质子(H+)泵等。主动运输在维持细胞内环境的稳定及功能上起着很重要的作用。
离子通道
由于现代光谱学、生物信息学和蛋白质结晶技术的发展,研究了离子通道蛋白的三维结构,从而对其转递过程有了较深入的了解。离子通道简单的理解就是离子通道蛋白在多种构象之间的转换,从而达到“开”与“闭”的两种状态。一般开与闭有扎门控制,称“扎门控通道”,极少数无扎门调控称“非扎门控通道”。扎门控通道可分三种:
- 配体门控通道:这些通道实际是离子通道型受体,它们与细胞外特定的配体结合后,发生构象变化,结果使“孔”打开,离子通透。典型的是乙酰胆碱酯酶受体及一些神经递质受体。
- 电压门控通道:膜两侧跨膜电位改变是控制通道的开关的直接因素。虽然跨膜电位只是以毫伏计,但膜通道感受的电场强度很大。由于这类通道的结构中有对电位改变的敏感区,所以可诱发整个通道功能状态的改变。这类通道一般是离子(K+、Na+、Ca2+、Cl-)通道。
- 机械门控通道:也是跨膜信号转换,既是外来机械信号,引起细胞跨膜电位变化。如内耳毛细胞顶部听毛受到切合力的作用产生弯曲时,毛细胞出现感受器电位,以传导信号。
离子通道受胞内外环境中多种物质的调控:如阳离子(H+、Zn+、Ca2+、Mg2+等)、代谢产物(ATP、ADP、脂肪酸)、胞内第二信使、神经递质、激素和一些气体(CO、NO)等。这些物质以不同的方式直接或间接地调节门控通道的状态。
囊泡运转
一、内质网与高尔基体之间的运转
细胞内膜系统之间的物质转递常通过囊泡运转方式进行。囊泡运转是一种高度有组织的定向运转,各种物质能够准确地运到靶细胞器,主要是囊泡的外面有各种不同的蛋白质包被,而在细胞器的胞质表面又有特殊的标志蛋白,不同的标志蛋白的组合,决定膜表面的识别特征。
囊泡外面的蛋白质有多种多样,可归纳为三大类:笼形蛋白、外被体Ⅰ及外被体Ⅱ。它们的组成及功能见表:
囊泡组成与功能
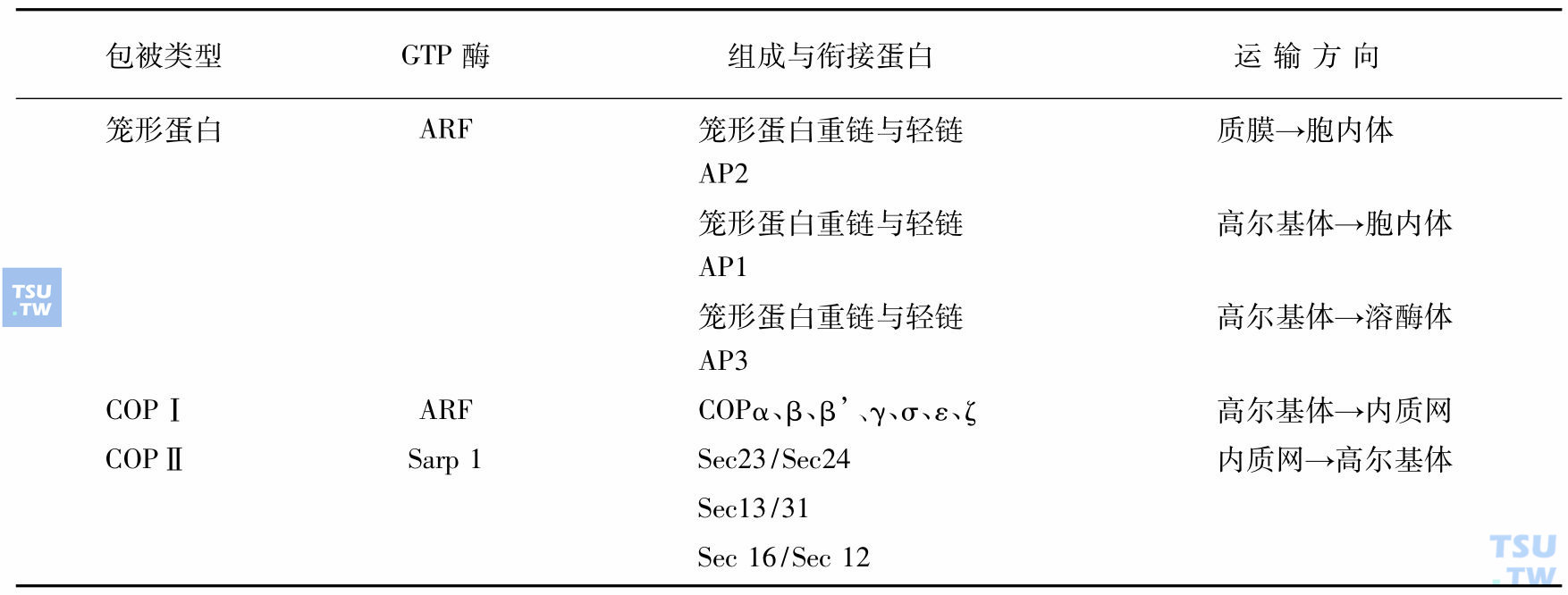
注:ARF:ADP-ribosylation factor,核糖基化作用因子。胞质内可溶性GTP结合蛋白,主要参与包被组装,对囊泡起调控作用。AP:adaptin,衔接蛋白;COP:coatomer,外被体;Sarp 1:GTP结合蛋白,具有GTP酶活性,在囊泡形成中起重要作用;Sec:Secretion从酵母中提出的蛋白,有分泌作用;Endosome:胞内体。指一系列与膜运转有关,形态及组成各异的膜系统的通称。
笼形蛋白分子是由3个重链和3个轻链组成,形成一个具有三个曲臂的形状,许多曲臂交织在一起,形成6边形网孔的笼子。笼形蛋白形成的包被中还有衔接蛋白(adaptin,AP),它介于笼形蛋白与配体受体复合物之间。现知衔接蛋白有三种:AP1、 AP2、AP3,可分别结合不同类型的受体,形成不同性质的运转囊泡。
COP原从高尔基体的囊泡中发现的,现分两大类:COP 1具有7种,按一定的比例在胞质中以复合体的形式存在。COPⅡ来源于酵母Sec蛋白家族,Sec23/24、Sec13/31分别与Sar1P(GTP酶)结合,它们包被在囊泡外,在囊泡形成过程中起主要作用。所以称为外被体(coatomer)。
二、分泌型囊泡与胞吐
胞吐作用即是胞吞的反方向。细胞通过胞吐作用分泌各种物质,被分泌的物质,有的吸附在细胞表面,另一部分成为细胞外液基质的成分,还有一些分泌到管腔或进入血液。细胞分泌途径有两种类型:固有型及调节型。
- 固有分泌:分泌物质持续不断地从高尔基体,经运输小泡送到细胞表面,通过胞吐作用将物质分泌到细胞外,一般指腺体的分泌,如胰岛素从胰岛分泌。分泌活动是一个非常复杂的循环过程。包括运转、定向、栓系、启动、触发、融合、胞吐等步骤。在循环过程的每个步骤都有许多复杂的调节因子参与。
- 调节型是蛋白质存在囊泡内,不立即释放,需等细胞接受信号后,才将蛋白释放。如内分泌激素及神经递质都属于这种类型。
能量代谢
线粒体是有机体能量代谢的中心,通过其内膜上的电子传递链氧化与磷酸化反应的耦联,生成ATP,是生命活动的能源。如线粒体受损,膜上一系列的氧化还原酶不能进行正常反应,能量不能产生,细胞立即死亡。特别是心肌细胞缺氧时,线粒体损伤,不能产生能量,心肌细胞即会很快死亡。
受体与信息传递
刺激与反应是一切生命现象的基础,而刺激与反应之间的耦联都依赖信息传递。如激素、神经递质、生物活性物质等,都是由一个细胞(或一组细胞)的信息传给另一个细胞。但有些物质是大分子化合物,不能直接穿过细胞膜,需要膜上的物质带入细胞内。膜上的这个物质即称“受体(receptors)”,受体必须有三个条件:
- 识别和结合:受体的某一部分的立体构象具有高度选择性,对某些配体(配体是指细胞外物质,又称第一信使)能识别并特异结合,这种结合是可逆的。单一细胞可能存在有多种类型的受体。
- 转导信号:受体与活性物质结合后,将信息传到细胞内,通过细胞内一系列反应,将所得的信息增强、分化、整合,并传递给后继效应机制(称第二信使)。
- 产生相应的生理效应:受体蛋白多是膜糖蛋白或糖脂蛋白,大都是镶嵌在膜内。受体种类很多,各自有其自己的反应通路,各受体通路之间又相互连接,形成一个大的网络,所以一个信息可能有多条通路,错综复杂。
根据受体的结构及效应体系的不同,可把受体分成3类:
- 门控离子通道受体:多由若干相同或不同的亚基组成,这些亚基围绕膜上的“孔道”排布,受体激活,使离子通道发生构相变化,孔道开放,导致离子跨膜流动,从而产生生物效应。一些神经递质的受体,如烟碱型乙酰胆碱受体(nACHR)、GABA受体等都是以这种方式发挥作用的。
- 与G蛋白耦联的受体:这种受体与配体结合后,立即与膜上的另一种蛋白耦联,使其释放出活性因子,活性因子再与效应器(指腺苷酸环化酶、CGMP磷酸二酯酶、磷脂酶C或离子通道等)发生反应。这种耦联蛋白都属一个家族,由于它能结合并水解GTP,所以称G蛋白。G蛋白种类繁多,但它们无论在结构和功能上都有许多共性。G蛋白是膜蛋白,由三个不同的亚基组成(α、β、γ),不同G蛋白主要是α亚基的差异。正因为有了α亚基的多样化,G蛋白才能实现对多种功能的调节。这类受体多是多肽类激素受体及神经递质受体等。
- 有酪氨酸激酶活性的受体:这种受体本身当它被激活之后,自身的酪氨酸残基磷酸化,从而产生生理效应。如胰岛素受体、某些生长因子受体、转铁蛋白受体及低密度脂蛋白受体等,都属这类受体。受体虽各自有自己的通路,它们之间互相促进又相互制约,以调节各信息之间的平衡。细胞膜上的受体也与其他蛋白一样,有合成及分解代谢。细胞膜上的受体数目一般都保持在一定水平内,但也受许多因素的影响,最主要的是配体本身的浓度,浓度愈高受体数目愈少,以这种自身调节,来维持正常的反应速度。有些疾病是由于受体数目的减少或受体的结构有变异而致病。
细胞表面抗原
血型特异抗原
人有ABO血型,A、B血型都有其特异的抗原性。决定血型抗原的是糖脂的糖。A与B血型之间的不同只差一个糖基,A型是乙酰半乳糖胺,而B型是半乳糖,O型是两种糖都没有,称O型血是万能输血源,即是这个原理。最近报道制出无血型的血液,实质就是将半乳糖或乙酰半乳糖胺除掉,形成O型血即可。
异体移植与组织相容抗原
排异反应是指一个人接受另一个人的组织或器官的时候,接受者对供者产生的抵抗,最终组织或器官脱离。现知排异的原因是由于异体组织或器官移植后,受者体内的T淋巴细胞产生一种抗体,释放到血液循环中,这种抗体是对供者细胞表面的抗原,在补体存在下,受者的血清抗体把供者的组织溶解。这种含有抗体的血清称“同种异型抗血清”。供者细胞(组织细胞或血细胞)上的表面抗原称“同种异型抗原”。由于这种抗原第一个发现在白细胞上所以称“人白细胞抗原(human leukocyte antigen,HLA)”。
这种相容抗原,在人及动物的组织及血细胞上都有,它是由一组紧密连锁的基因表达的分子,这组基因称“组织相容性复合体(major histocompatibility complex,MHC)”,有高度多态性区,可表达有多种异型。现知HLA有四类:HLA-A、HLA-B、HLA-C、HLA-D。HLA-D又分为HLA-DP、HLA-DR、HLA-DQ三种。
细胞识别
细胞的识别是指细胞对异物的识别及细胞与细胞之间的识别。细胞对异物的识别,如吞噬细胞吞噬细菌、病毒、衰老及变异的红细胞;又如病毒进入体内的第一步是连接到寄主细胞的表面,这种结合不是随机的,而是寄主体细胞质膜上的特殊部位,如红细胞有流感病毒的结合部位(血型糖蛋白A,glycophorin);疟原虫感染的部位。细菌、寄生虫及立克次体等在寄主细胞膜上都有结合部位,只是感染的机制不同。如细菌一般是它的毒素与细胞膜结合,结合后毒素进入细胞产生毒害效应。疟原虫进入红细胞是它生活史的一个阶段。
黏附分子(adhesion molecules)是涉及细胞与细胞、细胞与细胞外基质间的相互识别、相互作用的一类表面大分子受体蛋白。它分布于细胞表面和细胞外基质成分之中。通过与相应配体特异结合,参与细胞信号传导与活化、细胞的生长与分化、肿瘤的转移等一系列的生理病理过程。特别是造血系统,如骨髓造血干细胞的增殖分化,不仅需要生长因子,还需黏附分子黏附到基质细胞。血细胞的功能很多需有黏附分子的调节,如炎症反应中,白细胞黏附于血管内皮细胞,穿过血管壁,向血管外浸润的过程;血小板在受损血管壁上的黏附、伸展和聚集的过程;这些都是血细胞与血管内皮细胞膜上黏附蛋白之间的相互识别、介导的结果。
细胞膜黏附蛋白可分三个大家族:整合素(integrin)家族、免疫球蛋白(immunoglobulin)家族、选择素(selectin)家族及尚未归类的黏附分子。
整合素
是一组介导细胞黏附的细胞表面糖蛋白(受体蛋白)这些蛋白是由两个亚单位α和β以非共价键1:1结合的复合体。其分布及功能见表:
血细胞的整合素
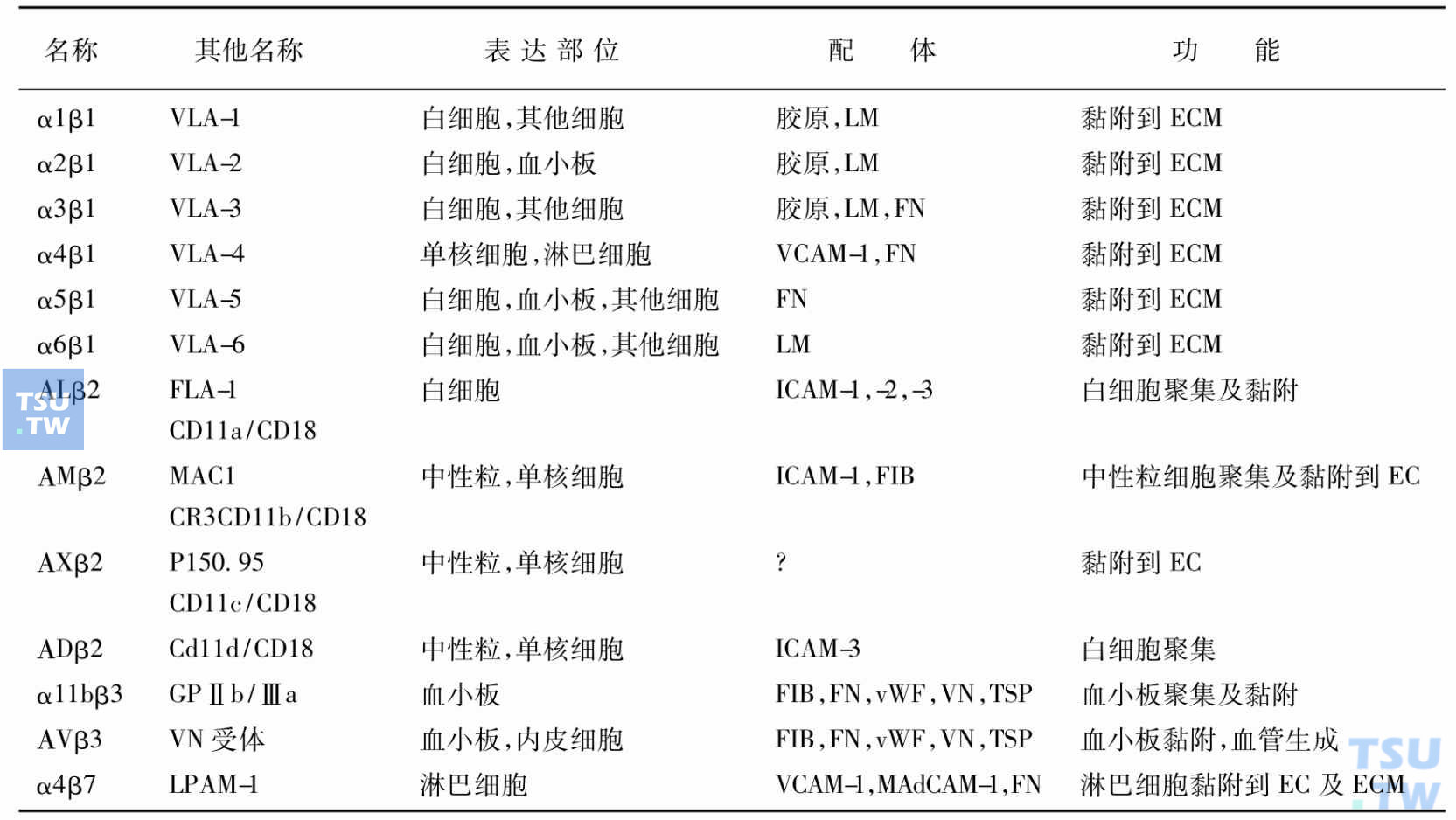
注:CR:补体受体;EC:内皮细胞;ECM:细胞外基质;FIB:纤维蛋白原;FN:纤连蛋白;GP:糖蛋白;LFA-1:白细胞功能相关抗原;LM:层连蛋白;LPAM:淋巴细胞Peyer黏附分子;MAdCAM-1:黏地址素细胞黏附分子;TSP:凝血酶敏感蛋白;VCAM-1:血管细胞黏附蛋白;VN:玻连蛋白;vWF:vW因子。
免疫球蛋白超家族
在结构上都有一个特点是通过二硫键形成环状,类似Ig功能区的结构。家族成员及其配体和功能见表:
免疫球蛋白超家族
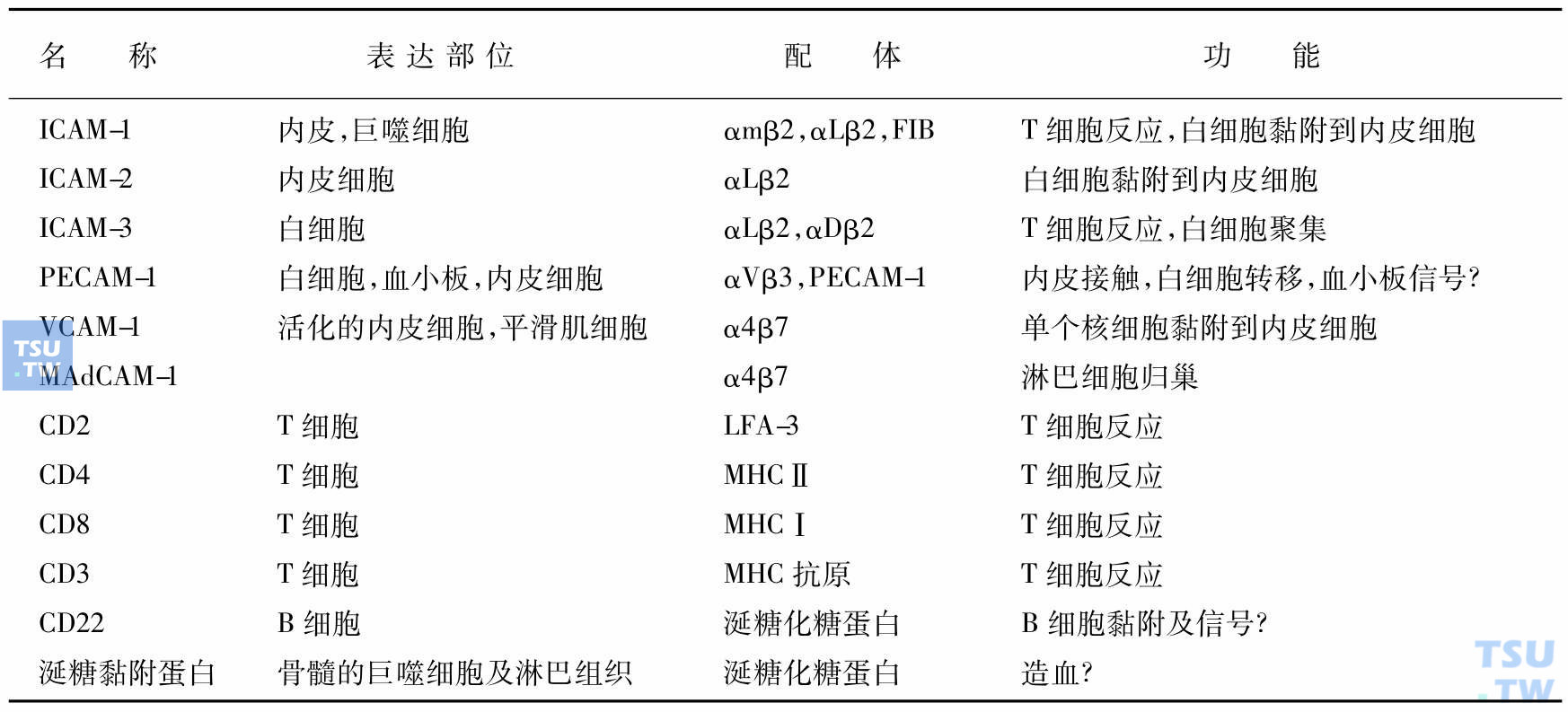
注:ICAM-1,-2,-3:细胞内黏附分子;PECAM-1:血小板及内皮黏附分子;MHC:主要组织相容性复合体。
选择素
表达在血管内皮细胞及白细胞表面,它们调节血管内皮细胞与白细胞的相互作用。由于这些蛋白都通过其外源凝集区域与靶细胞上的寡糖决定簇作用,选择性地促进细胞与细胞互相作用,因此称选择素。家族成员及其配体和功能见表:
选择素的表达部位及功能

注:ELAM-1:内皮的白细胞黏附分子;ESL-1:E选择素配体-1;GlyCAM-1:糖化的黏附分子;GMP-140:颗粒膜蛋白140;LAM-1:白细胞黏附分子;LECAM-1:白细胞内皮黏附因子-1;PADGEM:活化的血小板颗粒外膜蛋白;PSGL-1:P选择素糖蛋白配体。
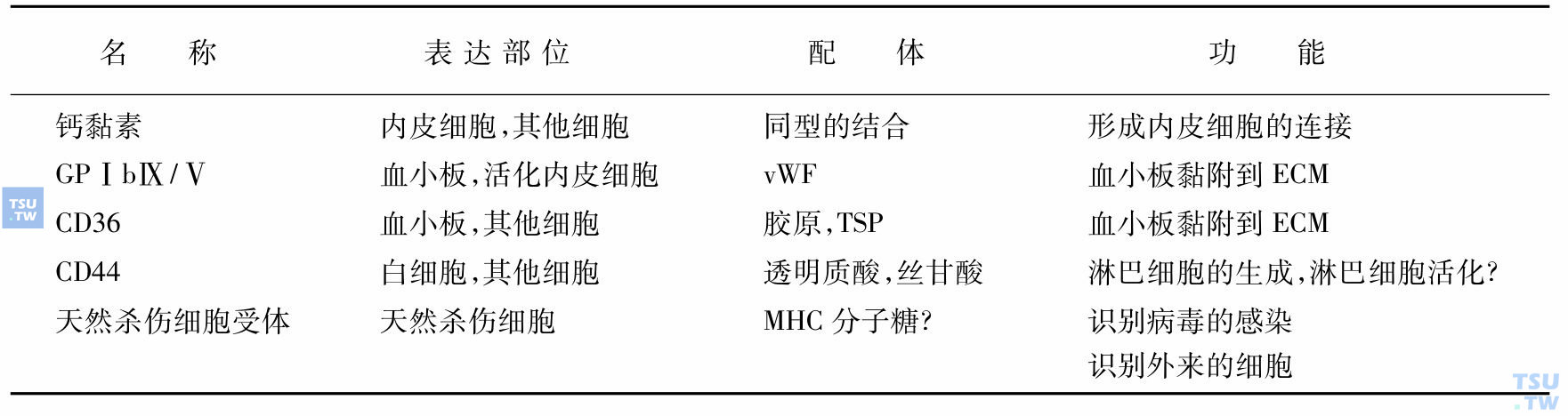
钙黏素(cahderin)其他黏附分子
介导细胞-细胞的接触,通过钙黏素将邻近同类细胞与细胞的连接。它与其他未归类的黏附分子的配体及功能见表:
钙黏素及其他黏附分子