
红细胞膜在红细胞生活过程中起重要作用,除维持红细胞的正常形态外,红细胞与外界环境所发生的一切联系和反应(如氧的传送、物质运输、免疫反应、信息传递以及药物作用等)都必须通过红细胞膜。
物质运输
细胞内外物质交换必须通过膜,红细胞内外气体、无机离子、糖、氨基酸等物质的浓度差别较大,许多物质的运转都有其各自的机制。
一、水的运输
膜脂是疏水的,水分子很难通过,所以它与离子一样需要有水的通道。水通道蛋白已克隆成功,称AQP(aquaporin)-CHIP。后来发现有7种AQP,红细胞上的称AQP1,从氨基酸的序列看,属于MIP (major intrinsic protein)家族。这个家族氨基酸的序列特点是有多个Asp-Pro-Ala。红细胞依赖水通道维持细胞内外的平衡,保持红细胞不被破溶。
二、葡萄糖运转
红细胞葡萄糖运转有葡萄糖运转体(glucose transporter),称GLUT,这个家族共有5种(GLUT 1~5),GLUT结构特点是它的C端及N端都伸向胞质面,跨膜部分穿膜12次。红细胞存在的是GLUT 1,其运转方式与阴离子通道相似,通过变构将葡萄糖从胞外运到胞内。
三、阴离子运转
红细胞阴离子运转主要是带3蛋白。其运转过程不需能量,但与细胞代谢有关,它主要介导HCO3-与Cl-进行1:1交换,以维持体内酸碱平衡。
四、阳离子运转
红细胞膜内外阳离子浓度差别很大,如胞外钙离子浓度是胞内的1000倍,它们的运输方式主要是依赖各种ATP酶的主动运输:红细胞膜上的Na+-K+-ATP酶可将胞内的Na+泵到胞外,同时把胞外的K+泵到胞内,所以又称其为Na/K泵,红细胞内钾含量相当于血浆中钾的30倍。Na+-K+-ATP酶由四个亚基组成,大亚基为120kD,小亚基55kD,依赖它们的变构将离子运转;Ca2+-Mg2+-ATP酶的作用是将胞内Ca2+泵出胞外,所以也称为Ca泵。红细胞膜中Ca泵的活性是Na/K泵的3~8倍,所以Ca泵在维持细胞内外离子浓度中起着重要作用。除以上这些ATP酶外,还有Na+-Na+、Ca2+-K+-ATP酶等进行离子交换。
受体
细胞外的信息物质要对细胞内产生反应都要通过与细胞膜上(或胞质中)相应的受体结合后引发一系列反应,这一过程即为信息传递,现知红细胞膜上至少有四类受体:第一类为激素受体(如胰岛素受体);第二类为递质受体(如去甲肾上腺受体);第三类为丙种球蛋白受体,如血型抗原可与相应的抗体结合;第四类为病毒(或细菌、寄生虫等)受体(或称结合点),如流感病毒感染。疟原虫裂殖子入侵红细胞,都是与红细胞膜上的血型糖蛋白特异结合,若将血型糖蛋白糖链上的唾液酸去除(或封闭),即不受感染。除这些受体以外红细胞膜上还有两个特异的受体。
红细胞生成素受体(EPO receptor)
EPO受体是由两条肽链组成,一个100kD,一个85kD,都有糖基化,经氨基酸分析两者很相似。它是跨膜蛋白,C端伸向胞内,N端在胞外,膜外部分有两个糖基化部位及多个半胱氨酸,最近发现EPO受体结构中约有200个氨基酸与白介素-2,3,4,5,6,7及G-CSF,GM-CSF的受体相同,共称为受体大家族(hematopoietin receptor superfamily)。这些受体N端的4个半胱氨酸形成两个二硫键。C端是Gly-X-Try-Ser-X-Try-Ser-X-Pro(X是可变的)。最近又发现其胞质部分与干扰素受体极为相似。用125I标记EPO受体,在人的CFU-E上约有1000个受体,其中有200个亲和常数是0. 1nmol/L(高亲和),其余的是7nmol/L(低亲和)。在进行反应时,可能都起反应,低亲和的是上调(up-regulation),高亲和是下调(down regulation),两者交替作用,以维持EPO的正常作用。EPO受体数目也随细胞发育阶段而异,在BFU-E阶段,受体开始形成,CFU-E时达最高潮,到早幼及成熟红细胞阶段逐渐减少,以至消失。EPO特异的作用在CFU-E阶段,当干细胞培养到CFU-E阶段,加入EPO,EPO与受体作用后,使Ca2+通道开启,胞内Ca2+浓度增高。3小时后发现有染色体结构变化,这时可能产生一种激活因子,促进珠蛋白基因转录,6小时才出现珠蛋白基因转录。转铁蛋白受体数20万~40万个/细胞。12小时开始血红蛋白合成,同时有带3及4. 1蛋白合成,12小时后细胞核脱落,成为成熟红细胞。
转铁蛋白受体(ferritin receptor)
体内血红蛋白代谢释放出的铁,在血浆内与血浆蛋白结合,形成转铁蛋白(ferritin),随血液循环运转周身,最早在胎盘及网质红细胞膜上提取到转铁蛋白受体,但成熟红细胞膜上无此受体。
转铁蛋白受体是跨膜糖蛋白,有两个同型的亚基,以二硫键相连,分子量为180kD,每个亚基结合一个转铁蛋白,一个转铁蛋白带两个铁,所以每个转铁蛋白受体带四个铁。人的转铁蛋白受体,N端62个氨基酸伸向胞质,C端672个氨基酸,膜外部分有648个氨基酸,插入膜有28个氨基酸。转铁蛋白与受体的亲和力很强,其次序为:带2个铁的转铁蛋白>带一个铁的转铁蛋白>载脂蛋白铁。正常人血浆含量50μmol/L。转铁蛋白受体有重要功能,似是吞饮的信号,当转铁蛋白与受体结合后,以吞饮方式转入细胞内,供血红蛋白的合成。
免疫功能
1981年有人提出“红细胞免疫系统”的概念,认为红细胞可清除免疫复合物(immune complex,IC),防止IC在组织沉积。从此,红细胞免疫研究迅速发展,大量研究表明红细胞不仅参与机体的免疫反应,还参与免疫调控,红细胞的某些免疫功能是其他免疫细胞所不能替代的。所以输红细胞也可以增强机体的免疫功能。
一、清除免疫复合物(IC)的作用
红细胞膜上有C3b受体(又称Ⅰ型补体受体,CR1),CR1和补体的作用是红细胞具有免疫功能的重要因素。由于红细胞数量多,血液中95%的CR1位于红细胞上,因此红细胞清除IC的机会比白细胞多500~1000倍。IC在周围组织的沉积是导致许多免疫性疾病的主要原因;另外如果过多的IC黏附在吞噬细胞等免疫细胞上,则会削弱它们的免疫功能。红细胞与IC的结合减少了IC对组织细胞的损伤;红细胞竞争性地黏附IC,可消除IC对吞噬细胞、淋巴细胞等免疫细胞的抑制作用,间接提高了它们的免疫功能。
二、对淋巴细胞的调控作用
红细胞能将IC结合的补体降解为C3dg,C3dg与红细胞膜上的CR2(Ⅱ型补体受体)结合,可诱导B淋巴细胞由静止期转向有丝分裂期,促使其增殖分化,并产生抗体。红细胞膜上的LEA-3(淋巴细胞功能抗原3)可与T淋巴细胞CD2作用,从而激活T淋巴细胞的免疫功能。另外,红细胞还能直接增强NK细胞的抗肿瘤作用。
三、对吞噬细胞的作用
红细胞膜上的CR1、CR3(Ⅲ型补体受体)可与吞噬细胞上的CR1、FCR(Fc受体)、CR3和CR4(Ⅳ型补体受体)共同作用,从而明显促进吞噬细胞的吞噬功能。另外,吞噬细胞在吞噬过程中释放大量的氧自由基,可对吞噬细胞造成损伤,红细胞上的超氧化物歧化酶(SOD)能够及时清除氧自由基,对吞噬细胞具有保护作用。
四、对补体活性的调节
补体包括20多种蛋白组分,当抗原抗体反应之后,经”瀑布式反应“最终形成补体的复合物,使细胞破溶,在补体一系列反应中有激活剂参与,也有抑制补体活化的分子参与,以调节补体的作用。红细胞膜上有三种抑制补体的分子:C3转化酶衰变加速因子(DAF,CD55),是针对C3转化酶的作用而命名的,它下调C3转化酶的活性,使C3不能转化为C3b,从而补体反应不能进行;反应性溶血的膜抑制剂(membrane inhibitor reactive lysis,MIRL,CD59),它主要抑制C9与C5b-8复合物的形成或抑制C9多聚化,抑制对膜的攻击。补体8结合蛋白(C8 binging protein),它阻止C9的聚合及膜复合体的攻击。这些因子在结构上都有一个共同点,虽然都是膜蛋白,但它们是含糖肌醇磷脂的蛋白,以磷脂的两个脂肪酸插入膜,蛋白质在膜外,由于它以肌醇磷脂插入膜,所以又有糖肌醇磷脂锚固蛋白(glycosylphosphatidylinositol anchored protein)之称。
五、氧化作用的清除剂
红细胞虽受氧及氮的侵袭,血红蛋白自身还产生自由基,血液长期渗透在病态的环境下(高脂、高糖、异物代谢产物),但红细胞内有大量的还原作用;如脱氧的血红蛋白通过亚硝酸还原酶作用可将亚硝酸盐还原成一氧化氮。当血红蛋白氧化成高铁血红蛋白,可通过高铁血红蛋白还原酶、五碳糖代谢产生的NADH、谷胱甘肽、谷胱甘肽还原酶系统使高铁血红蛋白还原成氧合血红蛋白。除此之外还有强的抗氧化剂:如维生素C、E,以及抗氧化酶,如超氧歧化酶(SOD)、硫氧还原蛋白过氧化物酶(thioredoxin peroxidase)、过氧化还原酶(perxiredoxin)。过去认为红细胞是氧化的场所,其实红细胞数量大,它有很多的抗氧化剂,清除氧化能力很强。
血型抗原
红细胞膜的抗原物质是由遗传基因决定的,其化学组成为糖蛋白或糖脂。在红细胞系统中,已发现400多种抗原物质,分属20多个血型系统。近来发现许多红细胞膜上的蛋白与血型抗原有关。对这些分子的结构与功能关系的研究,可能对血型抗原与疾病的关系有进一步的认识。现将它们的基因编码及功能列于表:
人类红细胞血型系统
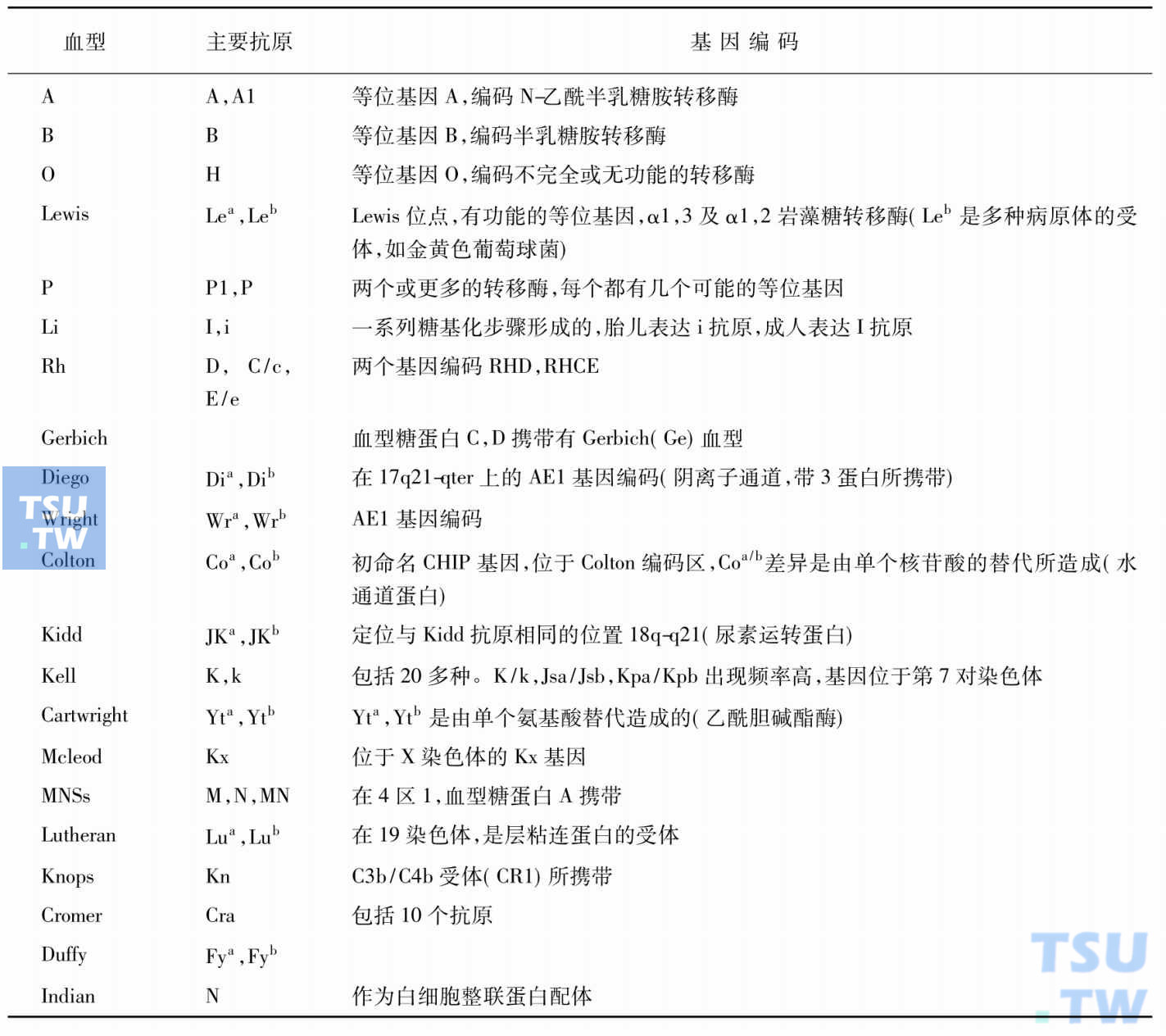
一、ABH抗原
红细胞ABH抗原不仅在血细胞上,在组织上也有,确切的称为组织-血型抗原(histo-blood group antigen)。ABH抗原存在两种形式:溶解型及膜型。溶解的抗原多存于内分泌或外分泌,如唾液、乳、尿及粪中;膜型的抗原存在血管上皮细胞、表皮细胞等膜上。溶解及膜上的抗原结构稍有差异。
二、Lewis抗原
Lewis抗原有两种:Lea及Leb,有三种表型:Le抗原是一种鞘糖脂。临床上发现Leb抗原是消化性溃疡的主要病因。
三、P抗原
P抗原包括三个主要的鞘磷脂抗原(Pk,P,PI):这些抗原的前体都是Gal(β1~4)Glc-ceramide。
最近发现多种寡糖抗原是各种感染源的受体。最引人注目的是P抗原是B19微小病毒及大肠杆菌的受体。B19小病毒是儿童时期常见的致病原,不表达P抗原的成人,血清检查证实无受过该病毒感染,体外实验发现红细胞的祖细胞亦不能被这种病毒感染。
四、I及i血型抗原
I及i抗原都是单糖或二糖。它们的不同只是L抗原多个支链。新生儿血内含i抗原较多,成人含I抗原,很少人有i抗原。可能N-乙酰葡糖胺转移酶在发育过程中调节。特异对I抗原的抗体表现出冷型自家免疫性溶血,这种患者血内含有抗体的滴度很高。体外实验抗体可结合红细胞并固化补体。正常人抗I抗体滴度很低。有些感染的疾病如单核细胞增多症及支源体肺炎产生冷凝集素,分别产生抗i及抗I抗体。
五、Rh蛋白和抗原
Rh D抗原是最具免疫源性的非ABO血型抗原。虽然已广泛使用Rh免疫球蛋白进行预防治疗,但Rh D一直是新生儿溶血的最常见病因。Rh血型系统包括50多种抗原,其中5种抗原(D抗原和C/c,E/e)认识得较清楚。这5种抗原被RHD和RHCE两个基因所编码,表达出至少两种或更多的蛋白。Rh系统内的多形性可能有两种机制:E和e抗原的表达,是由于单个核苷酸替代而导致的一个氨基酸的差异;而C和c抗原的表达则关系到4个核苷酸及氨基酸的替代。其他Rh多形性是由于产生新的基因结构,例如RHD或RHCE基因中的一个或多个外显子替代了其他基因的外显子。然而就个体而言,Rh抗原在什么条件下才表达仍未阐明。
Rhnull综合征是与异常红细胞和溶血性贫血相联系的Rh无效表型,患者缺乏所有的Rh抗原,其红细胞呈口形。有轻度的溶血性贫血。现知这类患者是因为RH50,一个编码Rh相关的基因位点,发生了突变。缺乏所有Rh蛋白的个体呈现出一系列复合的红细胞的异常,包括:口形红细胞、异常的阳离子转运、膜胆固醇的相对缺乏和非Rh相关的血型抗原(包括Lw、Ss、U和Fy5)的表达缺陷。带有D抗原的蛋白或是带有C/c和E/e抗原的蛋白,都是维持红细胞正常的形态和生存时间所需的。
六、Gerbich抗原和血型糖蛋白C和D
在Gerbich血型系统中有一种无效的表型(称作Leach)与椭圆形红细胞和极轻度的溶血性贫血相关联。携带有Gerbich抗原的蛋白现已证明为血型糖蛋白C和D(glycophorin C,D),二者都是血型糖蛋白位点上的产物。血型糖蛋白C是从第一个ATG密码子开始转录而形成的,而血型糖蛋白D则是自下游的框内ATG开始转录而合成的。血型糖蛋白C和D在膜蛋白中含量较少,可与带4. 1蛋白结合,是构成细胞骨架的主要膜蛋白。因此,血型糖蛋白C和D的变异型若在胞质内的一段是完整的,仍可与4. 1蛋白结合,则红细胞形态正常,反之若缺乏血型糖蛋白C和D,或无4. 1蛋白结合区的变异型,会导致4. 1蛋白的部分缺失出现椭圆形红细胞。
七、MNSs抗原和血型糖蛋白A及B
MNSs抗原与红细胞膜血型蛋白A及B有关。MN抗原在血型糖蛋白A上,有131个氨基酸,MN之间只有两个氨基酸不同。
血型糖蛋白A及B含有16个O-连接的糖苷键和一个N-糖苷键,但MN抗原不在糖链上,在蛋白部分。抗MN抗体是IgG及IgM的混合体,产生溶血情况极少。
Ss抗原在血型糖蛋白B上,A与B之间由于mRNA剪切的部位不同,B比A少70个氨基酸,前26个氨基酸是一样的,Ss两个等位基因只在29位氨基酸有变化,S是Met,而s是Thr。抗S及抗s抗体即是识别这两个氨基酸。
八、Kx蛋白和McLeod表型
Kx蛋白由X染色体上的一个基因所编码。这种蛋白的cDNA已克隆,并发现它编码的蛋白结构上与众多神经递质转运蛋白类似,而且携带Kx抗原。Kx蛋白对携带于另一个蛋白的Kell血型抗原的正常表达是必需的。完全缺失Kx蛋白的个体会出现棘形红细胞,继而出现神经症状。
九、蛋白衰变加速因子(DAF)与Cromer抗原
DAF的组织分布很广,它是一种糖肌醇磷脂跨膜蛋白,与一般膜蛋白不同,以脂肪酸插入脂双层。DAF可加速C3和C5转化酶的衰败,不论是来自经典或是替代补体激活途径都有作用。因此,DAF有抑制补体激活逐步放大的功能。
DAF携带有Cromer血型系统的抗原。这一系统包括7个出现频率高的和3个出现频率低的抗原。其中4种特异性重复区(consensus repeat domain)携带的抗原已确定:Cr(a-)、UMC、Tc(a-)和Dr(a-)。Inab表型的主要特征是完全缺失DAF和所有的Cromer抗原。Inab表型个体的细胞在体外对补体激发的溶血敏感度轻度升高,但这一现象在体内无重要意义。这种表型与疾病的关系,虽有两例Inab个体患有炎性或恶性的肠道疾病的报道,并没有明确的联系。出现Dr(a-)表型与部分DAF缺失和Dra抗原丢失有关。这些是由于一个核苷酸碱基被替代,和大部分未成熟的mRNA剪切错误,导致终止密码子提前出现,因此缺少与糖肌醇磷脂结合的部位,所以不能与膜结合,但这种表型对补体不敏感。
十、Duffy(Fy)血型抗原与细胞因子
Duffy血型抗原Fya和Fyb由第一对染色体FY位点上的等位基因产生。Fy抗原有以下几种现象:①许多非洲人和非洲裔美国人带有Fy(a-b-)表型;②Fy(a-b-)表型可抵抗间日疟原虫和Knowlesi疟原虫的感染。最近报道Fy抗原位于一个有多个跨膜区域的蛋白上。这种蛋白可结合多种细胞因子,包括IL-8、RANTES(调节激活正常T细胞表达和分泌)、黑色素瘤生长刺激活素和单核细胞趋化蛋白-1。特别是这些趋化因子属于C-C和C-X-C型趋化因子。而IL-8是疟原虫的竞争性抑制剂。Fya和Fyb抗原只因单个氨基酸的差异而不同,但并不影响它们与趋化因子或疟原虫的结合。然而Fy抗原在生理上的重要性仍不清楚。非洲起源的Fy(ab-)表型看起来是FY前体的GAA-1结合部位发生突变造成的。这一突变导致Fy蛋白在造血干细胞中不能表达,但在其他组织中仍可正常表达。迄今,由于编码区突变而出现FY基因无功能的家系只有一例报道。因此,设想FY基因对人的健康不是必需的。
十一、Indian(In)抗原与细胞外基质分子(CD44)
CD44糖蛋白是整合蛋白,广泛分布于组织中。编码CD44的基因是一个复合的基因,能编码许多剪接型,但成熟红细胞主要表达一种由外显子1~5 和15~19编码的剪接型。CD44是人类主要的透明质酸多糖(hyaluronan)受体,结合纤连蛋白、层粘连蛋白和骨桥蛋白(oesteopontin)。Indian抗原N-末端域与软骨连接蛋白同源,其分子结构仅有一个氨基酸的差异。红细胞CD44的表达是受到显性抑制性基因In(Lu)的调控,In基因还下调Lutheran抗原的表达。至今In(Lu)基因的表达产物仍不知道,也未能确定它是怎样调节这两种在基因和结构上均不相同的蛋白。许多组织中都观察到,CD44异构体的出现与分化有相关性。CD44的过度表达和它的某些剪接型的表达增加,似与很多肿瘤的恶变和转移倾向有关。
虽然早期工作认为红细胞上的CD44不具功能,近来的工作显示,CD44对纤连蛋白与早期红细胞的祖细胞连接很重要,而且CD44在镰形红细胞可能与纤连蛋白、层粘连蛋白的结合增多。有人提出CD44可能与AnWj抗原有关,后者是一种抗原表位,可作为流感杆菌凝血株和脊髓灰质炎病毒的受体。
十二、LW抗原与细胞黏附分子
LW抗原有LWa和LWb由一个42kD的糖蛋白所携带。这个蛋白与细胞间黏附分子家族同源,这一家族还包括白细胞黏附素(leucocyte adhesins)ICAM-1、ICAM-2和ICAM-3。LWa/b的多态性是由于单个碱基对的替代所造成的。LW(a-b-)是由于LW基因第一外显子内的一个碱基缺失,导致终止密码子提前出现而成。
十三、带3蛋白(下称带3)与Diego抗原
红细胞带3蛋白,又称作AE1,是红细胞主要的阴离子转运子或称阴离子交换蛋白。它对协助HCO3-和Cl-的速交换及CO2的运转很重要。AE1的分布很局限,在红细胞和肾表达最多。它属于一类多基因家族编码的蛋白。Diego血型系统有两个相应的抗原,Dia及Dib,Dia/b的多态性是由AE1蛋白结构中多次穿膜最后的一个环的一个氨基酸(脯氨酸或亮氨酸)多态性决定的。Dia仅在东南亚及印第安人中,其他人群很少见,Dib比较多。但Di (a-b+),Di(a+b-)表型的红细胞都能正常运转阴离子。
十四、水通道蛋白(aquaporin-1)与Colton (Co)抗原
红细胞水通道蛋白起初命名为CHIP。首先在纯化Rh蛋白时发现的。因为CHIP在大小上与Rh蛋白相似,也具有多个跨膜区域。CHIP现改名为aquaporin-1(AQP1),由于发现一例无AQP1表达的个体,同时也不能表达Colton血型抗原,从而发现AQP1与Colton血型的关系。AQP1基因位于编码Colton血型抗原的基因域。Colton抗原有两型:Coa 与Cob,抗原的差异是由单个核苷酸的替代造成的。最近发现在3个无亲属关系的Co(a-b-)表型个体中,其突变各不相同。这种表型的分子基础可能是异源性的。AQP1承担红细胞>85%的渗透水的转运功能,这些个体中并没有因缺乏这种蛋白而产生临床症状。Co(a-b-)者红细胞形态正常,但精确测量其表面积略有减少,生存期限稍微减低。对一例特殊的先天性红细胞造血紊乱性贫血的病例观察,患者的多种红细胞膜蛋白表达异常,包括AQP1和CD44的缺乏和血红蛋白开关失调,但患者的AQP1基因序列是正常的。
十五、Kell血型与内源多肽酶
Kell血型系统很复杂,至少有21个不同抗原,其中最主要的是K/k、Jsa/Jsb和Kpa/Kpb。其中K、Jsb、Kpb出现频率高。Kell抗原是糖蛋白存于膜内,抗原性很高,表达IgG型抗体,可产生迟发性输血反应及新生儿溶血,一般临床上很少,因为90%人是K (-)。Kell有临床意义。Kell抗原糖蛋白与内源多肽酶高度同源,可使激素失活,如脑啡肽、催产素、血管紧张素Ⅰ和Ⅱ、缓激肽等。另外,Kell抗原还与其他膜蛋白及红细胞骨架起作用。Kell抗原的表达依赖另一个37kD蛋白,它携有Kk抗原。Kk抗原基因在X染色体短臂近X-连接的慢性肉芽肿病及肌营养不良症的部位。
十六、Cartwright(Yt)抗原与乙酰胆碱酯酶
Yt抗原有两个型:Yta,Ytb。两型只差一个氨基酸(His-Asp)。由于发现可用抗Y抗体提纯乙酰胆碱酯酶,证实乙酰胆碱酯酶与Yt抗原的相关。Yta/ Ytb的多态性是由单个氨基酸替代造成的,这并不影响酶的功能。
变形能力
红细胞在外力作用下变形的能力,称红细胞变形性(deformability)。红细胞具有很强的变形能力,不然它难以生存。这是由于红细胞直径约为8μm,当其要通过直径只有2~3μm的脾窦毛细血管时,必然受到挤压,此时红细胞能改变形态,从盘状变为细长条状,因此得以顺利通过而不影响其正常功能。影响红细胞变形性的主要因素有:
- 膜骨架蛋白组分和功能状态:红细胞形态主要靠骨架蛋白之间的相互作用形成规则的网架,并依赖磷酸化与脱磷酸化的调节,使其随生理的需要变形。如果骨架蛋白某些组分短缺,或骨架蛋白各组分之间以及骨架与质膜之间的结合有缺陷,都可造成变形性降低。
- 膜脂质流动性:流动性大有利于变形。流动性变化取决于膜脂质组分的改变,特别是胆固醇/磷脂比值,若比值升高,流动性下降,变形性则相应减少。
- 细胞表面积与细胞体积的比值:正常红细胞呈双凹盘状,有较大的比值,变形性良好;若红细胞趋于口形或球形,比值减少,则变形性降低。
- 血红蛋白(Hb)的质和量:Hb在红细胞内占有很大的比重,它的变化会直接影响红细胞的形态。红细胞内Hb浓度增高,或变性Hb附着于膜上,均使膜变得僵硬,变形性降低。
- 膜的离子通透性:一般离子通过膜的速度很慢,相比之下极性弱的易透过,极性强的不易透过,例如,钾离子半交换期超过30小时,氯离子只需0. 2秒。在某些病理情况下红细胞的通透性发生改变,使Na+、K+通透性增加。如果Na+进入细胞量远大于K+的外漏,细胞内则积水,细胞肿胀;相反,若K+的漏出多于Na+的内流,细胞会脱水,体积减小,细胞内黏度显著增高。以上两种情况都会导致红细胞变形性降低。
维持出/凝血的平衡
红细胞对出血与凝血的作用很少有人注意。早在1910年Duke曾报道过,血小板减少的患者,输血后虽然血小板数未增加,但出血时间有改进,他提出红细胞可能对凝血有作用。50年后Hellen经大量的实验证实红细胞确与凝血有关。发现红细胞膜有缺陷(球形红细胞增多症)及血红蛋白异常(地中海贫血)的患者都有血栓的形成。他认为是由于红细胞的集聚及对内皮细胞的黏附性增加,促进血栓形成。体外实验观察,如加红细胞到血小板中,促进了血小板对玻璃壁的黏附性,在电镜下可观察到它们的集聚。Valles提出红细胞的游离脂肪酸及类花生酸代谢产物诱导血小板活化。另外,红细胞可促使血小板脱颗粒,释放ADP、5羟色胺、β血小板球蛋白,从而血小板集聚。他们还认为红细胞有多种隐蔽的黏附因子,正常生理情况下,是非黏附性的,当病态下(如球形、口形、地中海、镰性贫血)转化成黏附性的。这些因子如:CD44(透明质酸受体),LFA-3 (leucocyte function associated antigen-3),LW血型抗原,CD47(整联蛋白结合的及血小板反应蛋白(thrombospondin)受体,lutheran血型抗原(层粘连蛋白受体)和CD36(血小板反应蛋白受体,在成熟红细胞没有,在网织红细胞中含少量)。红细胞膜上还有爬行酶,它是维持磷脂双层不对称性重要的酶,当出血时,细胞内Ca2+增加,使爬行酶活化,促使丝氨酸磷脂(PS)外翻,PS是凝血因子的活化剂,可引起凝血的瀑布反应。
红细胞除了与凝血有关外,还有抗凝血的作用。体内在无外来刺激时,出/凝血机制处于动态平衡,当有微小变化时,可随时加以纠正。当血红蛋白脱氧时,伴有血红蛋白的结构转换,促使血红蛋白的β93半胱氨酸释放NO,NO有使微血管扩张及抑制血小板活化的作用。同样,在pH低、PO2低、剪切力低时,也可活化内皮细胞的嘌呤能受体(purinergic receptor),也促进NO产生。红细胞还可表达外酶(ectoenzyme),使ADP降解成AMP,可避免ADP对血小板的激活。
从以上红细胞在出/凝血中的作用看来,红细胞在维持机体正常生理状态下出/凝血的平衡起着一定的作用。
