
珠蛋白基因呈簇状分布,α-及α-类基因定位于第16号染色体的短臂,而β-及β-类基因定位于第11号染色体的短臂。α-类基因簇总长超过40kb,而β-类基因簇长度约60kb。
α类:16p13,基因簇(24Kb):5'—ζ—Ψζ—Ψα1—α2—α1—3'
β类:11p15,基因簇(60Kb):5'—ε—Gγ—Aγ—Ψβ—δ—β—3'
α-珠蛋白基因簇共包含七个连锁的α类基因或假基因。这七个基因的核苷酸顺序均已测定,并且除θ基因外,它们的功能状况也都已清楚。在α基因簇,有两个α-珠蛋白结构基因α1及α2位于复合物的右面,即3'端。这两个α基因均含三个外显子,被两个内含子顺序隔开。两个α基因在第二内含子区顺序有些区别。在α-表达基因的5'端或上游是一假的α-基因(Ψα),其DNA顺序密切地相似于α-基因,但有一个使之失去功能的突变。在其上游是两个胚胎期的α样基因(ζ-基因、Ψζ-基因),它们中的一个(Ψζ)无表达作用。
β-基因复合物从5'到3'端由6个基因组成:一个胚胎期基因(ε),两个胎儿基因(Gγ及Aγ),一个假β-基因(Ψβ),δ-基因及β-基因。这两个γ-基因产生多肽链,但两基因有差别,即在136位是甘氨酸的是Gγ,而系丙氨酸者则为Aγ。由于δ-及β-基因之间有广泛的同源性,因而认为δ-及β-基因来自单个基因的复制。δ-基因的调节区域接着突变,致使其功能无意义。上述α-及β-珠蛋白基因簇在染色体上按顺序排列基因,在胚胎期、胎儿期和出生后发育过程中,它们被表达。这些基因中每一个及基因内部DNA的核苷酸序列都已进行过检测。
所有功能珠蛋白基因的结构普遍相同,由三个外显子(为珠蛋白链密码顺序)及两个内含子或间插序列(intervening sequence)(此序列并不翻译成蛋白质)组成。下图显示了珠蛋白基因的结构。其中,5'及3'非翻译区DNA序列对基因表达是很重要的。“启动子区”由紧接转录开始的起点处约100个碱基对组成。在此区内,有三个结合催化mRNA合成的RNA聚合酶的短序列。有两个序列:TATA盒及CAT盒,对基因转录的起始特别重要。该序列发生突变作用则会使其与酶的结合力降低,从而限制mRNA的转录。第三个外显子下游,含有序列:AATAAA,它是基因转录的终止信号。现在认为该区域能触发酶的作用,可以在适当的位点切断并释放mRNA,进行进一步的加工。
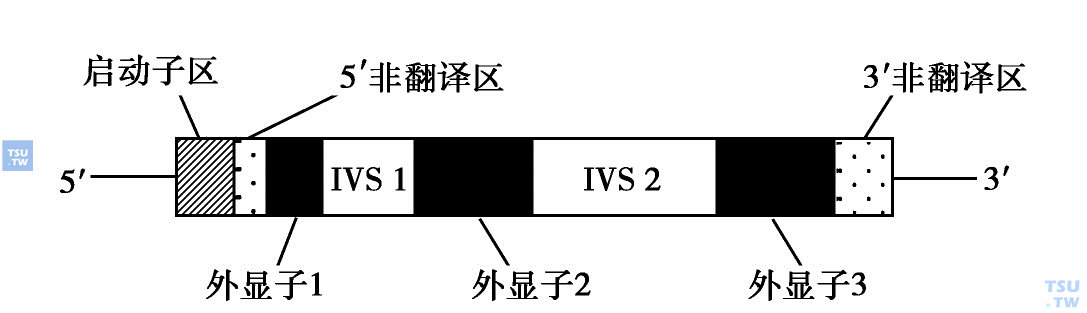
珠蛋白基因的结构:每个人珠蛋白基因含有3个编码部分(外显子),其被2个内含子分开。在β-珠蛋白基因中外显子1编码第1~30个氨基酸,外显子2编码第31~104个氨基酸,外显子3编码第105~146个氨基酸。阴影部分为5'及3'转录、非翻译区。启动子区约含100bp,其中有些是转录起始所必需的。
mRNA转录子包含完整的基因组DNA序列拷贝,即外显子与内含子,编码146个氨基酸。在该mRNA前体被运送到胞质之前,要经过5'端带帽,剪切去除由内含子转录的序列及3'端多聚腺苷酸化,这是个加工过程。最后一步对于完成产物由核到胞质的运送以及mRNA在胞质中的稳定性是非常重要的。剪切过程中外显子末端的下游(供体位点)与下一个外显子末端的上游(受体位点)相接近使前体mRNA形成环状结构。然后酶解切除内含子序列,且封住外显子的供体与受体位点。几乎所有真核基因,每个内含子的5'端均有GT序列,3'端为AG序列。前者称为给位裂解信号(供体位点),后者称为受位裂解信号(受体位点)。已发现5'端和3'端切接位点所在的短序列(一般为9核苷酸)具有高度保守性,称为通用序列(consensus sequence)或“共有位点(consensus site)”,它们的通式分为5'……AG/gtagt……3'和5'……()nn ag/G……3(n >11,大写字母表示外显子部分,小写字母表示内含子部分)。突变作用引起剪接结合部位及共有区的异常剪接,因此产生异常mRNA。
