
中性粒细胞的吞噬作用
中性粒细胞到达感染源后即可通过各种受体识别经补体或抗体调理的细菌颗粒。C3b、C3bi和免疫球蛋白是非常重要的调理素,它们在细胞表面的相应受体分别是CR1、CR3和FcRⅠ、FcRⅡ、FcRⅢ。调理的细菌与受体接触并被中性粒细胞表面的黏附分子粘住、开始了吞噬作用;首先由伪足沿着与细菌结合的部位从四周伸展包抄,最后收口直至脱离质膜形成吞噬体。前已介绍中性粒细胞吞噬细菌,形成吞噬体的过程是一个由Ca2+及多种特殊功能蛋白调控的肌动蛋白、骨架网络结构变更的复杂过程,而且中性粒细胞功能行为的某一过程又为下一进程创造了条件,这种情况在吞噬作用与消化杀菌作用的衔接过程中更为明显。例如,当中性粒细胞的伪足在捕捉细菌进而形成吞噬体时一部分质膜内陷,使质膜上的NADPH氧化酶亚基之一的细胞色素b558与胞质内另一个氧化酶亚基胞质因子得以接触,并装配成活化的NADPH氧化酶,开始了氧依存性杀菌进程。同时,吞噬体形成,脱离质膜进入胞质后与中性粒细胞溶酶体颗粒接触并融合,引发了脱颗粒作用,开始非氧杀菌进程。
杀菌消化作用
吞噬体形成后几乎同时开始非氧杀菌和依氧杀菌进程,并且互相配合,协同完成杀菌及消化过程。
脱颗粒作用与非氧杀菌
随着吞噬作用的引发,中性粒细胞的脱颗粒作用也启动,中性粒细胞的颗粒移位并与质膜内陷形成的吞噬体融合,形成吞噬溶酶体或称消化泡。颗粒中的各种抗菌蛋白水解酶随即释放出来,此过程称脱颗粒作用。脱颗粒作用的发生标志着非氧杀菌作用的启动,也为依氧杀菌作准备。特异颗粒较初级颗粒更快地释放它们的内容物,但过量的内容物可能会通过不完整的吞噬泡而分泌到细胞外。初级的嗜天青蓝颗粒与溶酶体非常相似,因为它们含有几乎在中性粒细胞中能找到的各种酸性水解酶,而且吞噬体内的酸性环境为这些酶提供了适宜的pH环境,酸性条件也是进一步脱颗粒作用的基本条件;阴离子通道的闭锁,可削弱Na+和H+的流出,从而可以削弱中性粒细胞的脱颗粒作用,但颗粒的碱性化却可促进中性粒细胞的分泌作用。虽然杀菌作用和消化作用发生在消化泡内,但中性粒细胞的功能犹如一把双刃剑,因为颗粒内容物分泌到胞外后,会造成组织损伤。脱颗粒作用也可发生在中性粒细胞与组织的黏附表面,并随之与免疫复合体、聚集的免疫球蛋白、补体组分或某些细胞因子相结合。这种状况下,颗粒内容物的释放是通过与内吞作用相反的过程(逆内吞作用方式)或无效的吞噬作用进行;颗粒与质膜融合,它们的内容物直接排放到中性粒细胞外。
细胞骨架蛋白对脱颗粒作用有重要作用,这些蛋白促进颗粒向吞噬体或质膜转移,无论是微管还是微丝,均参与了此过程。中性粒细胞激活后三秒钟,颗粒就结合到聚集的微管上,抑制微管的聚集作用,从而阻止中性粒细胞的脱颗粒作用。肌动蛋白聚集犹如在质膜和颗粒之间筑起一道屏障,对颗粒分泌起抑制作用。中性粒细胞受到微粒或可溶性刺激物激活时,杀真菌剂代谢物如细胞松弛素B等,可强烈促进颗粒分泌,这是因为其破坏了胞质中肌动蛋白微丝,使颗粒膜和质膜易于融合。有关膜融合的精确机制目前尚不得知,但已知中性粒细胞中有若干个对Ca2+敏感的磷脂酶参与此过程,导致颗粒-吞噬体或者颗粒-质膜接点处的脂质组分改变,磷脂酶A2(PLA2)能促进组成上与质膜相似的颗粒与脂质体的融合,因为该酶降低了Ca2+浓度。PLA2存在于静息的中性粒细胞颗粒膜上,而且PLA2以分泌型的形式转位到吞噬体上。特异颗粒和分泌泡在低Ca2+浓度下,比初级颗粒更容易与人工脂质体[500μmol/L的磷脂酸/磷脂酰乙醇胺(1:3)]融合。当Ca2+浓度高至12mM时,后者的融合即终止。胆固醇加入到脂质体或颗粒制品中时,能增强融合,游离Ca2+对中性粒细胞初级颗粒、特异颗粒、三级颗粒的胞吐作用和分泌泡的形变是必需的条件。分泌泡对低浓度Ca2+敏感,相反,初级颗粒则需较高的Ca2+浓度。中性粒细胞颗粒对Ca2+浓度的这种巨大差异,在中性粒细胞渗出到皮肤窗口时,也能观察到。根据对中性粒细胞分泌液的分析,证明分泌泡的全部、38%的白明胶酶颗粒、特异颗粒中22%的乳铁蛋白可被动员,但仅7%的MPO可从初级颗粒中被动员。对有渗透性的中性粒细胞的研究表明,1μM(在生理范围内)的Ca2+即可导致特异颗粒的标志物CD67升调节,而对初级颗粒的标志物CD63升调节则需要GTP-γS和1μM的Ca2+同时存在。中性粒细胞的初级颗粒含有两种GTP结合蛋白(25和26kD),而特异颗粒和质膜则有一套三种分子量在20~23kD范围内的GTP结合蛋白、浓度为0. 3~1. 0mM的Mg-ATP和其他三核苷酸。对有渗透性的中性粒细胞也有激活脱颗粒作用,这就支持了有一个共有的三核苷酸受体调节中性粒细胞脱颗粒作用的观点。有两种腺苷酸受体位于中性粒细胞上,而且已证实腺苷可抑制脱颗粒作用和呼吸暴发作用。
刺激中性粒细胞可激活磷脂酶D,促使磷脂酰胆碱转换成磷脂酸。膜组分上的这种改变使一种融合性的磷脂取代了非融合性的磷脂,进而促使脱颗粒作用的发生。膜联蛋白在促进中性粒细胞与含有磷脂酸和磷脂酰乙醇胺的脂质体融合时,对磷脂酶浓度的依赖不如Ca2+。膜联蛋白Ⅰ和Ⅱ两种蛋白对促进磷脂囊泡与中性粒细胞膜的胞质面的融合作用有重要意义。经胰蛋白酶处理的中性粒细胞或者中性粒细胞膜制品,就不能再与富含融合性脂质的脂质体融合,这充分证明了包括膜联蛋白Ⅰ在内的胞质蛋白在中性粒细胞脱颗粒作用中起重要作用。另一些融合蛋白的同系物,如25kD的突触小体相关蛋白(SNAP-25),是神经末梢部位胞吐作用的主要驱动者。另一些位于中性粒细胞质膜上对突触小泡的停泊和融合起作用的蛋白是分泌泡相关膜蛋白-2 (VAMP-2)、突触融合蛋白-4和SCAMP,SCAMP蛋白在中性粒细胞的白明胶酶颗粒、特异颗粒、嗜天青蓝颗粒及分泌泡中也有发现。
促分泌剂是一种可溶性分子,它既可激活中性粒细胞也能促使颗粒内容物释放到胞外。理想的促分泌剂应是既能促使初级颗粒和次级颗粒释放内容物,又能引起Ca2+和cAMP浓度的瞬间上升。不完整的促分泌剂虽然能引起Ca2+的增高和释放特异颗粒作用,但没有改变cAMP的水平。包括C5a、fMLP、PAF、LTB4和Ca2+载体A23187等大多数趋化物都是完整的促分泌剂,而不完整的促分泌剂包括有PMA和细胞表面激活剂、ConA等。一些非趋化性的活性物质如白介素、细胞因子和髓系生长因子,其作用既像完全促分泌剂又像非完全促分泌剂。炎性介质TNF-α、IL-1和IL-3对用细胞松弛素-B处理的中性粒细胞均能使初级颗粒和次级颗粒释放。然而,IL-4和趋化性IL-8只对特异颗粒的释放作用有影响。GM-CSF可引起中性粒细胞释放特异颗粒,而且其在对用细胞松弛素-B处理的中性粒细胞作用时,很像一种完全的促分泌剂,单独GM-CSF(7. 5μg/kg)作用于正常人时,可引起除初级颗粒以外的所有颗粒表达β-2整联蛋白CD11b。GM-CSF还能诱导趋化性IL-8的释放,而对IL-3和TNF-α则不能。单独G-CSF(300μg)也能升调节CD11b及FcγRⅢ,而且导致胞质乳铁蛋白和弹性蛋白酶的增高。每天持续给予G-CSF可提高中性粒细胞的绝对计数量,而且明显提高了TNF-α水平,但对IL-8或GM-CSF胞质水平则无影响。虽然较微刺激中性粒细胞时,特异颗粒中的乳铁蛋白有50%,初级颗粒中MPO有90%的放出,但仍可能使三级颗粒白明胶酶全部耗竭,表明在这类中性粒细胞的低水平激活作用中,三级颗粒白明胶酶的显著性释放与中性粒细胞的始动是相关联的。
吞噬体形成后即与胞质中的溶酶体颗粒(初级或次级颗粒)发生融合,形成吞噬-溶酶体或称消化泡。颗粒中的各种抗菌蛋白水解酶随即释放出来,此过程称脱颗粒作用,开始了非氧杀菌消化进程。在消化功能上,溶酶体颗粒中的抗菌蛋白分子可分为杀菌性的和消化性的两种,如次级颗粒中的乳铁蛋白可螯合铁离子,夺取细菌生长必需的基本元素;同时乳铁蛋白与铁离子形成的复合物又是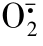 与H2O2通过Haber-weiss反应生成重要毒性氧物质羟基自由基OH.的催化剂。在初级颗粒中有一种阳离子蛋白能与带负电荷的真菌结合而抑制细菌生长,BPI则通过增加细菌质膜的通透性达到杀死细菌,就杀菌这点而言,非氧杀菌与依氧杀菌相比是次要的,只是依氧杀菌的一种补充。但杀菌后的消化作用则须依靠溶酶体中许多蛋白水解酶来完成;溶菌酶主要消化细胞内的糖肽,也水解某些腐生菌的细胞壁黏多糖。酸性水解酶是主要的消化降解酶,初级颗粒中的中性蛋白酶(组织蛋白酶G和弹性蛋白酶)更是大分子蛋白强有力的降解酶,在这些酶的协同作用下被杀死的细菌(或机体自身的死亡细胞)彻底被消化,消化泡剩余的膜则可被再利用,用以补充质膜,废弃物则通过泡吐作用排出胞外。
与H2O2通过Haber-weiss反应生成重要毒性氧物质羟基自由基OH.的催化剂。在初级颗粒中有一种阳离子蛋白能与带负电荷的真菌结合而抑制细菌生长,BPI则通过增加细菌质膜的通透性达到杀死细菌,就杀菌这点而言,非氧杀菌与依氧杀菌相比是次要的,只是依氧杀菌的一种补充。但杀菌后的消化作用则须依靠溶酶体中许多蛋白水解酶来完成;溶菌酶主要消化细胞内的糖肽,也水解某些腐生菌的细胞壁黏多糖。酸性水解酶是主要的消化降解酶,初级颗粒中的中性蛋白酶(组织蛋白酶G和弹性蛋白酶)更是大分子蛋白强有力的降解酶,在这些酶的协同作用下被杀死的细菌(或机体自身的死亡细胞)彻底被消化,消化泡剩余的膜则可被再利用,用以补充质膜,废弃物则通过泡吐作用排出胞外。
非氧杀菌的调控:
随着分子生物学学科的发展和分析技术水平的提高,下列颗粒内容物已在基因水平上被克隆,而且对其5'上游或旁侧序列也已判定。它们有MPO、弹性蛋白酶、组织蛋白酶G、蛋白酶3、溶菌酶、防御素、乳铁蛋白、gp91phox、p22phox、CD11b、CD18、维生素B12结合蛋白、hCAP-18/FALL-39、NGAL、白明胶酶及碱性磷酸酶。虽然已经知晓众多转录因子在启动域中可确定的结合位点,但实际上能确定相应启动域的只有极少数,如MPO、组织蛋白酶G、溶菌酶、弹性蛋白酶、CD11b、CD18、gp91phox和白明胶酶的转录因子。一些转录因子可能参与一些颗粒内容物表达的调控,如在乳铁蛋白,CD11b的基因上有GATA-1转录因子的结合位点,转录因子GATA-1存在于造血祖细胞的早期,促使细胞分化。在人GM-CSF,小鼠MPO和弹性蛋白酶基因上有AML(PEBP2/CBP)转录因子的结合位点。在蛋白酶3和嗜天素(azurocidin)基因上有转录因子C-MYB的结合位点。PU1转录因子对造血系统有特异性,而且在B淋巴细胞、粒细胞和单核细胞中有高表达。在粒细胞各个分化时期都有PU1表达。许多基因都含有PU1的结合位点。在粒细胞分化期间,PU1转录因子对某些颗粒内容物的转录开始较早,如弹性蛋白酶、GMCSF受体和G-CSF受体,而对某些颗粒蛋白则转录较晚,如CD11b、CD18和FcγRI等。另外在弹性蛋白酶、G-CSF和G-CSF受体基因上有C/EBP2转录因子的位点,而且证实C/EBP2经与其他转录因子如C-MYB(弹性蛋白酶)或PU1,很可能还有新的转录因子,被陆续发现参与颗粒内容物蛋白表达的调控。细胞骨架蛋白对脱颗粒作用有重要调节作用。细胞骨架促进颗粒向吞噬体或质膜转移,无论是微管或微丝,均参与此过程。中性粒细胞激活后三秒钟,颗粒就结合到聚集的微管上,抑制微管的聚集作用从而阻止中性粒细胞的脱颗粒作用。肌动蛋白聚集就如在质膜和颗粒间筑起一道屏障,对颗粒分泌起抑制作用。细胞松弛素B等可强烈促进颗粒分泌。腺苷也可抑制脱颗粒作用和呼吸暴发作用。
依氧杀菌机制
依氧性杀菌作用主要通过NADPH氧化酶的激活、利用O2大量生成超氧阴离子(),然后再形成一系列有强氧化作用的衍生物如H2O2、OCl-、OH-及氯胺等实现,这些物质统称活性氧物质(ROS),ROS破坏细菌的蛋白质分子、核酸及酶等重要生物分子而杀死细菌。依氧性杀菌过程由多个环节组成,包括磷酸己糖通路激活和NADPH氧化酶激活,氰化物不敏感性氧消耗激增和ROS大量生成,如下图所示。
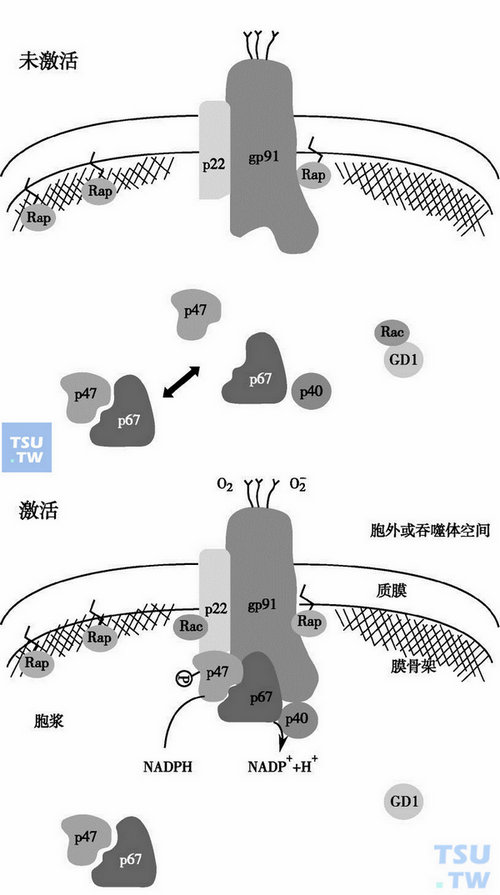
NADPH氧化酶的激活示意图
NADPH氧化酶是整个依氧杀菌机制的心脏,NADPH氧化酶是至少由八种分别位于中性粒细胞各个部分的蛋白组成的多组分酶系统(下表),统称吞噬细胞氧化酶(phagocyte oxidase,phox)。当调理性吞噬作用或促分泌作用激活中性粒细胞期间,这些蛋白的某些组分移位到皮层细胞骨架和吞噬体膜或质膜上。静息中性粒细胞中,p47phox与p67phox相结合,p67phox又与p40phox形成复合体,NADPH氧化酶的激活是由于PKC介导的p47phox蛋白磷酸化,随之p47phox磷蛋白与p67phoxp40phox复合体相结合,并移位到质膜与黄素细胞色素b558,一个由22kD的α亚基(p22phox)和91kD的糖蛋白(gp91phox)组成的异二聚体一起组成的NDAPH氧化酶。三种胞质组分的结合发生在中性粒细胞的皮质细胞骨架上,而且在该处p47phox被完全磷酸化。大约八成的细胞色素b558位于特异颗粒、白明胶酶颗粒和分泌泡中,并在中性粒细胞激活时移位到皮质细胞骨架和质膜。p47phox的磷酸是使与细胞骨架和膜相关的NADPH氧化酶组分从胞质转位至质膜并进行装配的基本要素。
低分子量GTP结合蛋白Rap1A是与黄素细胞色素b558紧密结合的蛋白,对NADPH氧化酶功能的完整性是必需的,Rap1A在质膜和特异颗粒膜上均有发现;而且在调理性吞噬杀菌时,与细胞色素b558一起移位到皮质细胞骨架或吞噬体膜,当Rap1A被磷酸化时,就脱离细胞色素b558。Rap1A的活化与呼吸暴发作用无关,而且慢性肉芽肿患者的中性粒细胞的Rap1A仍可正常活化。
NADPH吞噬氧化酶(phox)组分的性质
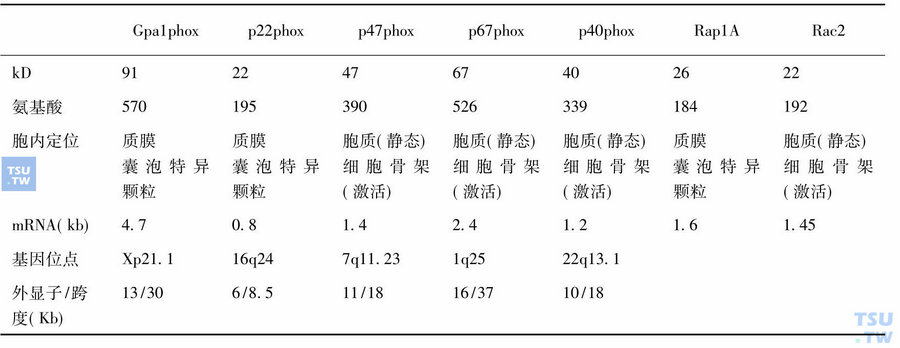
在中性粒细胞激活时,一种胞质GTP结合蛋白Rac2也与p47phox和p67phox-p40phox复合体一起被移位到细胞骨架,正常情况下,Rac1和Rac2在与GDP解离抑制因子Rho-GDI结合成复合体时,均处于不活化状态。Rap1A或Rac2对NADPH氧化酶活性均有调节作用,主要机制是它通过与相应底物GTP而非GDP的结合与解离而进行调节。中性粒细胞激活时,生物活性脂质例如花生四烯酸、磷脂酸和磷脂酰肌醇即被生成,这些活性脂质可以解除Rap1A与细胞色素b558的结合,和Rac2与GDI的结合而使Rac2被激活;Rac2的激活是p47phox和p67phox-p40phox复合体移位所必需的。活化的Rac2的作用犹如一种“穿梭蛋白”的作用,携带phox蛋白到皮层细胞骨架,然后Rac2再变成与GDP结合的非活化状态,返回到胞质。中性粒细胞活化时,p47phox的丝氨酸残基(Ser)被高度磷酸化,然而,研究发现只有p47phox C末端的第379位的Ser被磷酸化,表明该Ser对NADPH氧化酶的活性或p47phox与细胞骨架的结合起关键作用。在非细胞系统中,Rac蛋白对p47phox和p67phox的全部活性也是必不可少的。细胞色素b558是杀菌型超氧化物生成系统NADPH氧化酶复合体的终端组分,无论是其α还是β亚基均含有血红素基团,这种结构充分适应了电子从胞质NADPH到黄素蛋白再到细胞色素b558传递的需要,进而跨膜传递至吞噬体表面或细胞外表面,在该处O2被还原成,在O2成为O-•2的过程中,电势变化从-330mV分别变为-256mV至NADPH,-245mV至黄素蛋白,-160mV至细胞色素b。与线粒体内的细胞色素相比,这些中间电势相对较低,p67phox促动电子从NADPH向黄素蛋白流动,p47phox对电子从细胞色素b558中的黄素蛋白传递到血红素、加工成超氧化物是必不可少的,NADPH氧化酶由细胞色素b558和胞质因子两个亚基组成。细胞色素b558位于质膜上,又由分子量91kD和22kD的两个蛋白组成。胞质因子位于胞质或溶酶体膜上,由47kD(p47phox)和67kD (p67phox)组成。在未发生吞噬作用时,细胞色素b558和胞质因子因分布于不同区域而自然分离,当吞噬作用发生时,质膜的细胞色素b558和胞质因子蛋白因质膜内陷和溶酶体向质膜移动而相互接触,并自然组装成NADPH氧化酶复合体并因磷酸化作用而活化,氧化酶激活的梗概示意于图48-4中。
趋化因子刺激如何引起氧化酶组装或激活一直是依氧杀菌的核心问题。通过多年研究已经肯定蛋白激酶C(PKC)的激活是NADPH氧化酶激活的必要前提,这是基于如下重要实验事实:①用PKC激活剂佛波酯(PMA)激发中性粒细胞呼吸暴发产生超氧阴离子期间,发现其与胞质亚基p47phox和p67phox转移到质膜相关联,而且还发现p47phox在细胞骨架上重新分布;②测得p47phox和p67phox与酪氨酸激酶Src家族SH3或A微区同源结构域上有与骨架蛋白作用的位点;③用PKC抑制剂星形孢菌素(staulosporine)可以抑制p47phox磷酸化作用并可破坏PMA引起的超氧阴离子()产生。近年对NADPH氧化酶激活与蛋白激酶的相关性研究有了较快的进展,逐年又有新的蛋白激酶被发现参与趋化因子激活中性粒细胞的呼吸暴发,特别是发现酪氨酸激酶乃是整个刺激瀑布系统中最重要的调控系统。此后又发现丝/苏氨酸激酶也是很重要的与氧化酶磷酸化有关的激酶,至今已报告有酪氨酸激酶(TPK)、蛋白激酶C(PKC)、丝裂原活化蛋白激酶(MAPK)、Ca2+/钙调素依存性蛋白激酶Ⅱ(CAMPKⅡ)、S6-激酶Ⅱ和组蛋白H4蛋白激酶(PKH4)等多种蛋白激酶参与激活中性粒细胞的呼吸暴发作用。由于NADPH氧化酶的激活及调控非常复杂,而且还涉及胞内-核内信息传递及氧化应激产物的基因表达,因此对其本质的揭晓将是艰巨和令人激动的。
氧依存性杀菌在吞噬作用后数秒钟内即开始,此时中性粒细胞的氧耗率可提高100倍。磷酸己糖通路活性可增强10~20倍。NADPH氧化酶的功能是利用磷酸己糖通路中的电子使氧分子(O2)转变为超氧阴离子()。中性粒细胞大量耗氧生成的生理行为称呼吸暴发作用(respiratory burst)。

产生的本身仅呈微弱的杀伤性,但其衍生成的强氧化物具有极强的杀伤性。在超氧化物歧化酶(SOD)作用下通过下式可生成H2O2。
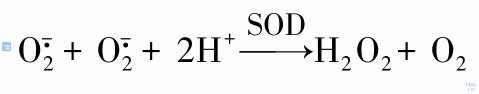
H2O2是很重要的氧代谢物,一方面其可以直接作用于细菌,还可以在髓过氧化物酶(MPO)作用下产生杀菌力更强的OCl-。反应式如下:
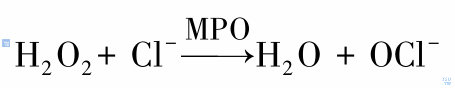
OCl-是极强的杀菌性氧化物,可与多种生物组分如血红蛋白、卟啉、硫醇蛋氨酸、还原型吡啶核苷酸、胺和氨基酸发生反应。而且其与胺反应可生成氯胺,也是极强的杀菌剂。
从上可知,自H2O2经MPO催化可以生成多种强杀菌作用的物质,故这个系统在中性粒细胞氧化杀菌中举足轻重,是中性粒细胞氧化杀菌的主力,常把此称作H2O2-MPO系统或简称MPO系统。Weiss等证明呼吸暴发时可测定的O2的40%是通过MPO系统产生杀菌性氧代谢物的。
虽然H2O2在Fe2+存在下还可与经过Weiss反应生成OH-也有较强的杀伤性。但OH-和ROO-等活性氧物质的生成是在某些特定条件下才出现的,而且维持时间较短,还不能与MPO系统的作用并论。由于H2O2-MPO系统中有H2O2、OCl-和氯胺等强杀菌物质产生,因而可以抵抗多种细菌、真菌和病毒等的侵害。
中性粒细胞内还存在一种防止H2O2过量生成、避免危害细胞自身的防护机构,即过氧化氢酶及谷胱甘肽循环系统,前者可直接分解H2O2,后者可将H2O2转变为无毒的H2O。尽管如此,中性粒细胞杀菌作用时产生的ROS还有分泌到细胞外的可能,加之MPO系统中的OCl-很易对抑制溶酶体释放的中性蛋白酶的α-抗胰蛋白酶造成损伤,失去对这些酶的抑制能力而造成周围组织损伤。因此,中性粒细胞的杀菌防御功能具有二重性,中性粒细胞氧化损伤是炎症及心、血管病变的重要原因。
激活的氧化酶复合体只需通过其亚基在平面上的简单分离又可回复到静息状态。
近已证明,NADPH氧化酶并不是唯一在质膜或吞噬体内组装并激活的,在特异颗粒中也可激活并产生氧化剂,并能保留在胞内。
