
CD8+淋巴细胞在刚刚离开胸腺时尚未分化为有功能的细胞。静止或天然CD8+T细胞是细胞毒细胞(CTL)的前体细胞。它们表达T细胞受体和CD8+分子,通过TCR和MHCⅠ类分子可以识别抗原。在与抗原相互作用后,CD8+T淋巴细胞分化成为CTL。
CTL细胞主要对同种异体细胞、病毒感染细胞、肿瘤细胞、化学物变性的细胞起作用。诱导CTL的抗原是在同种异体细胞上的Ⅰ型和Ⅱ型主要组织相容性抗原(MHC)。Ⅰ型抗原诱导能力更强。CTL是抗原特异性的,仅溶解诱发其产生的细胞。由Ⅰ型抗原和非MHC抗原诱导的CTL有所不同。CTL仅在诱导其生成的抗原结合在Ⅰ型分子上呈递给它们时,才能杀伤靶细胞。CTL的这种性质叫做MHCⅠ型限制性。生理条件下,CTL的产生需要CD4+ Th细胞的帮助,Th受刺激时产生细胞因子,帮助CD8+细胞转化成CTL。前体细胞到CTL的分化分段发生,需要两个信号,第一个特异性信号是由抗原通过TCR/CD3来传递,第二个非特异性的信号来自细胞因子。
最主要的细胞因子是IL- 2和IL- 12。在CTL分化中,来自抗原呈递巨噬细胞的细胞因子刺激CD4+Th细胞增殖。对Ⅰ型和Ⅱ型抗原刺激形成不同的应答细胞,前者产生抗原特异性CTL,而后者提供辅助细胞功能的增殖信号。两种类型MHC分子的作用对CTL的产生是必需的。通常检测CTL的分析用51Cr标记靶细胞。当靶细胞CTL杀伤标记的细胞后,51Cr释放到培养液,可以检测出放射活性。病毒感染时诱导CTL是缓慢的,需要4~7天才会产生。
CTL溶解靶细胞的机制
靶细胞溶解的机制不依赖于抗体或补体。CTL溶解靶细胞的第一阶段通过TCR的参与(识别),CTL与靶细胞建立联系,通过黏附分子而黏附到靶细胞上,其中最重要的黏附分子是LFA1。LFA1的配体是细胞内黏附分子1(ICAM1),在多种来源的细胞中广泛地表达。另一黏附系统是CD2/LFA3复合物。这些抗原非依赖黏附系统加强CTL/靶细胞结合物的形成,TCR提供特异性和效应物作用。
CTL与靶细胞接触时核移到一边,颗粒挨在与靶细胞黏附的位置,与膜融合,与靶细胞接触常在4分钟内完成。融合标志着第二阶段的开始,细胞内出现变化。变化中最突出的是微管组成中心(MTOC)与高尔基体一起重新定位在与靶细胞接触的位置。CTL杀伤作用是Ca2+依赖性的,当CTL的靶细胞是另一个CTL时(特别是一个无关的靶细胞),仅靶细胞被杀伤。在效应物CTL/靶细胞CTL结合物中,MTOC重新定位仅发生在效应CTL内。MTOC对接触面的定位可能对CTL传递高尔基体小泡中的溶解分子有用。第三阶段为传递致死信号。CTL介导的杀伤涉及两个主要机制,分为穿孔素依赖的和穿孔素非依赖的。穿孔素介导细胞溶解时,CTL功能必须依赖穿孔素。缺乏穿孔素时,CTL不能在体外溶解病毒感染的上皮、神经外胚层或中胚层起源的异源的靶细胞。NK细胞的细胞溶解活性也依赖于穿孔素。穿孔素通常存储在颗粒内。
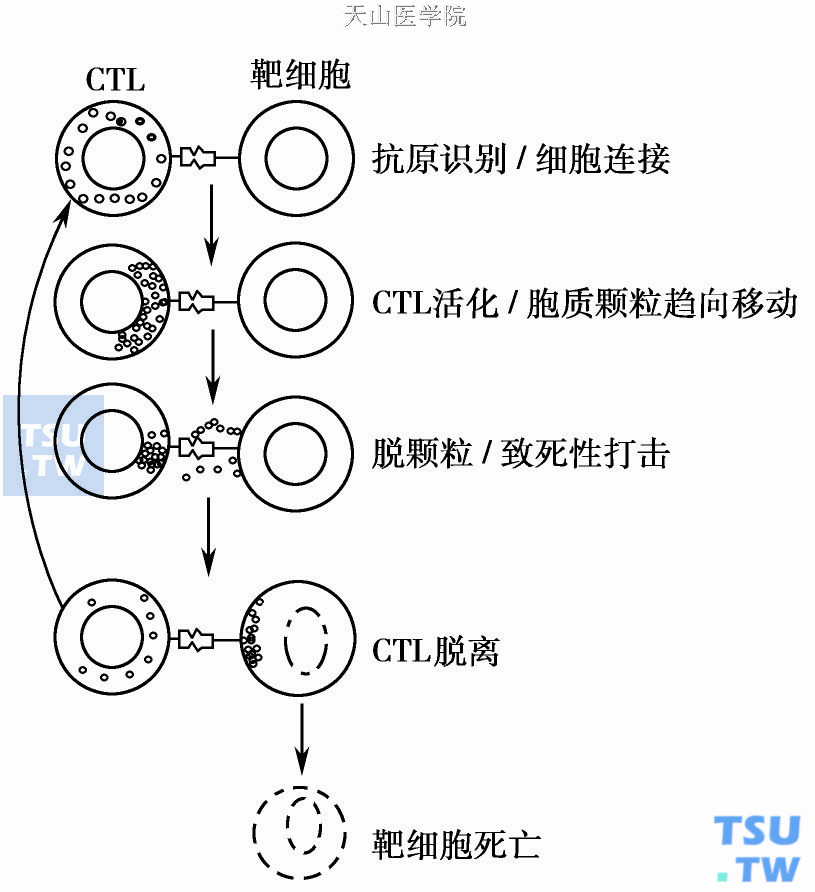
CTL攻击靶细胞的过程
细胞毒作用介导凋亡
从穿孔素基因的突变细胞发现,穿孔素是CTL介导的细胞毒作用的主要工具。然而,像肥大细胞瘤P815和淋巴瘤L1210这样的一些靶细胞,在缺乏穿孔素时仍然可被溶解,而且,靶细胞死亡涉及核变化,即染色质浓缩和DNA裂解,这是细胞凋亡的特征。有人认为CTL有不依赖穿孔素的机制。细胞凋亡依赖于Fas分子的表达,当Fas在靶细胞上出现时,介导CTL的细胞毒作用。穿孔素介导的细胞毒机制和Fas介导的细胞毒机制是两个特有的CTL细胞毒机制,与所有T细胞介导的细胞毒作用有关。
在CTL颗粒中鉴别出丝氨酸蛋白酶的一个家族称为粒酶。其三个常见的活性物是胰蛋白酶样成分,包括粒酶A、粒酶B和甲硫氨酸裂解酶(METase)。粒酶B是一种丝氨酸蛋白酶,粒酶B以非活性形式出现,必须切开N端肽的二肽基肽酶才激活。基因突变使粒酶B无效的小鼠模型中,同种异体反应的CTL不能诱导靶细胞的DNA裂解和凋亡。提示凋亡介导的细胞毒需要粒酶B。粒酶自身没有催化活性,在与一定量的穿孔素联合时,才能观察到靶细胞的DNA破坏。说明粒酶起作用一定要进入靶细胞内,进入途径可能通过由穿孔素形成的孔。这也说明为什么加载抑蛋白酶肽那样的蛋白裂解抑制剂时,会抑制靶细胞的杀伤。粒酶是丝氨酸蛋白酶,被认为从半胱氨酸蛋白酶发展来。线虫CED3基因编码这样的酶,与哺乳动物ICE是同源的。
粒酶B介导的凋亡会绕过由蛋白bcl- 2家族的凋亡抑制作用。bcl- 2可作用于ICE样蛋白酶的上游,抑制其活力。bcl- 2蛋白是线虫的CED9蛋白同源物,与CED3、CED4蛋白作用相反,在细胞存活中起维持作用。一种观点认为bcl- 2作用于调控ICE蛋白酶活性的蛋白。然而,bcl-2位于线粒体膜外层,有人认为它通过抑制线粒体PT阻止凋亡,因此抑制凋亡启动因子的释放。
靶细胞溶解末期发生一系列以产生大泡为特点的膜改变。在靶细胞大量释放51Cr前已观察到线粒体的破坏。最初的损害是仅允许小无机分子的交换,但进一步的更大的分子随之而丢失。细胞的最后破坏是由胶体渗透压不平衡引起的。然后,靶细胞的核起泡,被破坏,染色质浓缩,DNA破坏,细胞程序化死亡。这些改变可在细胞溶解期前检测到,应与发生在溶解后的DNA溶解区别开。在一些病理情况下,如大量病毒感染或大量肿瘤负荷,孔形成机制增加了CTL对靶细胞的杀伤性。一旦致死信息传递完成,CTL就移开去寻找另一个靶细胞。CTL如何与靶细胞分离的生化机制尚未得知。
