
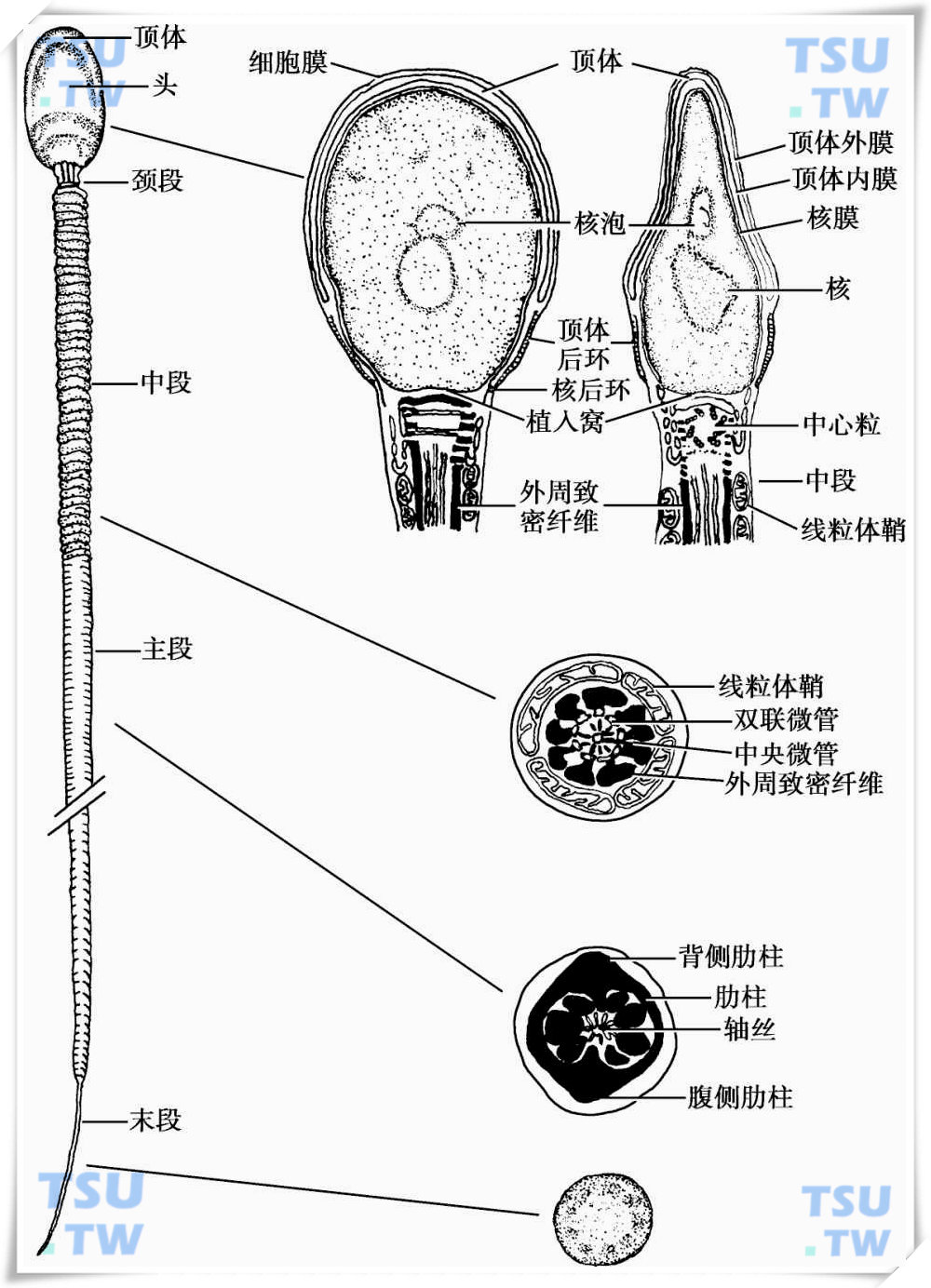
精子尾部又称鞭毛(flagellum),长约55μm,从头至尾可分为颈段、中段、主段和末段4个节段。
颈段
为精子头和尾的连接部位,故又称连接段(connecting piece),由前端的小头(capitulum)、后端的节柱(segmented column)和中央的中心粒组成(图1-1)。小头由致密纤维样结构组成,与核后端的植入窝相接触,两者间存有狭窄的间隙,其内含有细丝连于两侧的小头和核膜,起连接作用,如用胰酶消化细丝,则头、尾于此处断裂开来。节柱为小头向后延伸形成,首先形成2根较大的和5根较小的节柱,自前向后与尾部的长轴平行排列,此处的节柱呈节段状,每根节柱由9~10个节段相连组成,节柱的前端包围着近端中心粒,末端与中段的外周致密纤维相连。近端中心粒由9组双联微管组成,中心粒是节柱和轴丝形成的组织中心,精子尾部的微管由此长出延伸,并由此带动了精子的拉长。
中段
位于颈段和主段之间,长5~7μm,由内到外,主要由轴丝、外周致密纤维、线粒体鞘和细胞膜组成(上一篇《精子头部》的下图)。
人精子超微结构模式图
轴丝(axoneme)
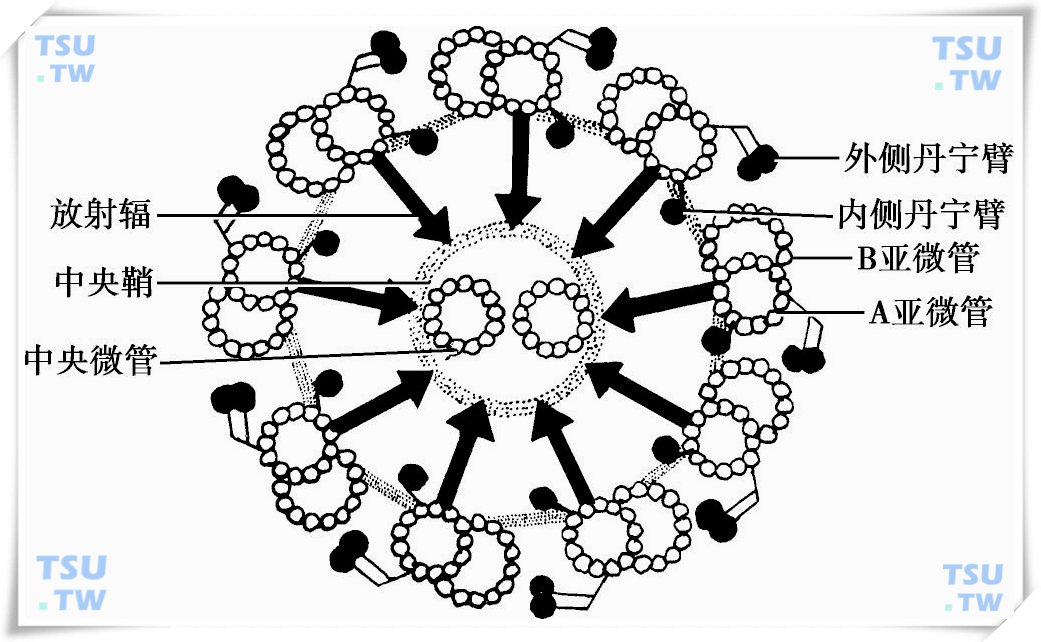
前端连于颈段,后部向尾部延伸,贯穿于精子尾部的全部,和纤毛的结构一样,轴丝由周边的9对双联微管和中央两根单独的微管组成,每对双联微管分为A亚微管和B亚微管,其中A亚微管管型完整,离中轴较近,B亚微管呈“C”字形,以其开口的两端附着于A亚微管。每根亚微管均由螺旋形排列的原丝组成,A亚微管有13根原丝,B亚微管有10~11根原丝围在A管一侧,另有3根原丝与A管合用。中央2根单独的微管管型完整,外包有中央鞘,根据双联微管的部位,将9组微管编为1~9号,编号原则为在横断面上,通过两个中央微管连线的中点作一垂直线,与此相交的一个双联微管为1号微管,然后按顺时针方向将其余8对双联微管分别命名为2~9号。每个A亚微管向下一个B亚微管伸出两个短臂,称为动力蛋白臂,分为内侧动力蛋白臂(Inner dynein arm,IDA)和外侧动力蛋白臂(Out dynein arm,ODA),主要由动力蛋白组成。
ODA与精子运动密切相关,用0.5mol/L的NaCl处理精子鞭毛,则精子的拍节频率降低了一半,ODA由多个多肽链组成,在人精子有两条重链(HC),每条450 000~530 000,2~3条中等大小链(IC),每条64 000~142 000,4~10条轻链(LC),每条8 000~30 000,ODA对维持精子鞭毛的运动方式并不重要,但影响到精子鞭毛拍击的力量和频率,研究认为,LC与精子运动密切相关,而IC和HC的功能并不清楚,用IC1的单克隆抗体作用后,精子的运动能力受到抑制,说明了IC的可能功能,IC69和IC78连在一起,与四条LC共同组成动力蛋白臂的基部,IC和LC形成的复合体可能与动力蛋白臂与下一微管的结合有关。IDA沿着轴丝由头到尾,IDA又分为三类,大约每96nm的距离为一种IDA,称为I1、I2和I3,I1由两个动力蛋白HC和两个IC(140 000和110 000)组成,研究表明,缺少IDA I1和I2时,精子仍能运动,说明IDA并不是精子运动必需的,但基因突变研究发现,ODA决定精子鞭毛摆动的速率,而IDA则能调节鞭毛摆动的节律。相邻的双联微管间有连接丝相连,长约20nm,自A亚微管的内、外支臂之间伸向B亚微管,连接丝的作用可能是控制动力蛋白与下一微管之间的间隙大小。
中央鞘向周围双联微管中的A亚微管发出丝状的结构,称放射辐,长80~100nm,将中央微管和双联微管连接起来(下图),放射辐呈杆状,一端连于A亚微管,另一端呈球状,称辐射丝头,指向中央微管。衣藻的放射辐杆部由17种蛋白质组成,其中杆部有12种,头部有5种,杆部和头部蛋白具有磷酸化。
人精子去细胞膜轴丝横断面模式图
微管的化学组成为蛋白质,主要的蛋白成分有:
一、微管蛋白(tubulin),相对分子质量为110 000,20℃时的沉降系数是4.8s,每个微管蛋白分子又由α-微管蛋白和β-微管蛋白两个亚单位组成,相对分子质量分别为55 000和60 000,是含谷氨酸和门冬氨酸较多的球形酸性蛋白。α-微管蛋白含450个氨基酸,β-微管蛋白含有445个氨基酸,二者间只有少数氨基酸残基不同。每个微管蛋白分子实际上是由2个亚基所组成的异二聚体,许多微管蛋白分子彼此首尾相接形成微管蛋白原丝,再由13条原丝排列组合成圆筒状的微管。中央微管和双联微管的微管蛋白略有不同,中央微管上有GTP的结合位点,而双联微管上则结合有ATP酶,微管蛋白与精子运动密切相关,抗微管蛋白抗体可抑制精子的运动能力。
二、ATP酶,又称动力蛋白(dynein),为双联微管支臂的主要成分,相对分子质量约500 000,为Mg2+依赖的ATP酶,酶活性占鞭毛ATP酶总活性的50%,其余的酶活性在放射辐的头部。动力蛋白的作用是分解ATP,每个动力蛋白分子每秒钟可分解11~35个ATP分子,精子鞭毛在一次摆动中能水解一个ATP分子,使化学能转变为机械力,使精子能够运动。体外ATP酶抑制剂可抑制精子的运动能力。
三、驱动蛋白(kinesin),相对分子质量约为600 000,由三个亚单位组成,可能参与微管的相互移动。
四、连接蛋白,又称为尼克辛蛋白(nexin),位于二联体之间的连接丝上,相对分子质量为150 000~165 000,起稳定轴丝、使其保持为一个整体的作用。
关于精子尾部能够摆动的机制,一般都接受“滑动微管模型”假设,鞭毛的运动不是微管收缩的结果,而是两个双联微管的A管和B管间可相互滑动,这主要是放射辐和中央鞘之间连接和脱离连续往返进行的结果,二联体臂上的ATP酶能水解ATP,产生能量,供鞭毛运动所需。微管结构异常,如周围微管或动力臂缺乏均可导致精子运动功能丧失,如纤毛不动综合征就是由动力臂缺失引起精子不能游动,同时体内的纤毛也不能摆动,使得在不育的同时伴有呼吸道疾病。
外周致密纤维(outer dense fiber)
围绕轴丝,由9根纵行的柱状结构组成,编为1~9号,分别与相同编号的双联微管靠近。外周致密纤维头端与颈段的节柱相连,尾段伸达主段长度的60%处。每根外周致密纤维的内侧与双联微管相邻,故将纤维给予与相邻双联微管相同的编号。不同部位的外周致密纤维粗细不一,起始段较粗,以后逐渐变细,1、5、6号纤维较其他的粗,横断面上,纤维呈梨形,底部朝外。致密纤维由蛋白质组成,半胱氨酸含量较高,蛋白质内含有大量的二硫键,因此较为稳定,外周致密纤维的作用主要是作为精子尾部的骨架,同时与精子尾部的弹性回缩有关。由于在精子鞭毛近端外周致密纤维较粗,鞭毛甩动时的弯曲振幅较小,随着外周致密纤维逐渐变细,局部抗弯力降低,弯曲振幅也相应增大。因此外周致密纤维的异常可导致精子运动方式的改变,如原地打转。人外周致密纤维是多基因的产物,其中一个基因编码蛋白为27 453 000(241个氨基酸),含二个外显子和一个内含子,定位于8号染色体上,基因突变可导致致密纤维排列异常。
线粒体鞘(mitochondrial sheath)
包绕外周致密纤维,由线粒体呈螺旋形包绕形成。人精子线粒体鞘有10~12圈,每个人精子约含75个线粒体,线粒体为椭圆形的细胞器,直径为0.5~1.0μm,每一线粒体由外膜、内膜及基质3部分组成。内膜又向内折叠形成平行排列的膜片状嵴。线粒体的主要功能是产生能量,在线粒体的内膜及基质中含有多个参与三羧酸循环的关键酶及能量转换的耦联磷酸化装置,为精子运动提供ATP。此外,线粒体还含有丰富的脂肪和磷脂,可维持膜的稳定性,需要时还可作为能量来源。线粒体数量减少或呼吸酶链缺陷均可导致精子活力下降。
终环(terminal ring)
在中段线粒体鞘最后一圈的尾侧,细胞膜反折特化,形成一层致密的板状结构,称终环。细胞膜附着于此环上,可防止线粒体在精子运动时向尾端移位。
主段
为精子尾部最长的一部分,长约45μm,组成精子尾部的主要部分。和中段相比,主段的轴心仍为轴丝,但外周致密纤维仅达头侧的60%。此外,主段无线粒体鞘,但在外周致密纤维的周围包有纤维鞘。
纤维鞘(fibrous sheath)由背侧纵柱、腹侧纵柱和环形肋柱组成,背侧和腹侧纵柱与精子长轴平行,分别起始于轴丝的第3和第8对双联微管的外侧。在主段的头侧,附着于第3和第8根外周致密纤维,在尾侧外周致密纤维消失后,两纵柱的内侧各发出纵行的嵴状突起,与相对应的双联微管相连。由于有9对双联微管,因此在横断面上纵柱将主段分成不对称的两部分,一部分含4根外周致密纤维(4,5,6,7),另一部分含3根(1,2,9)。肋柱是由紧密排列的环形细丝组成,两端变宽,与纵柱相连,肋柱之间的间隙较小,且相邻肋柱发出分支相连。纤维鞘的功能是调整精子尾部摆动的平面,由于3号和8号双联微管与背侧和腹侧纵柱相连,限制了微管的滑行运动,只有其余7对双联微管可以滑行,同时由于纵柱的存在使得精子尾部的弯曲只能发生在与背、腹轴相垂直的平面上,也就是在与中央微管连线相垂直的平面上。此外,纤维鞘上还含有三磷酸甘油脱氢酶,为糖酵解酶,提示纤维鞘还与精子的能量产生有关。
末段
为精子尾部的最后一段,起始部有少量纤维鞘,随着末段的变细,纤维鞘消失,仅剩中央的轴丝和外周的细胞膜,末端轴丝的双联微管可相互分离。
