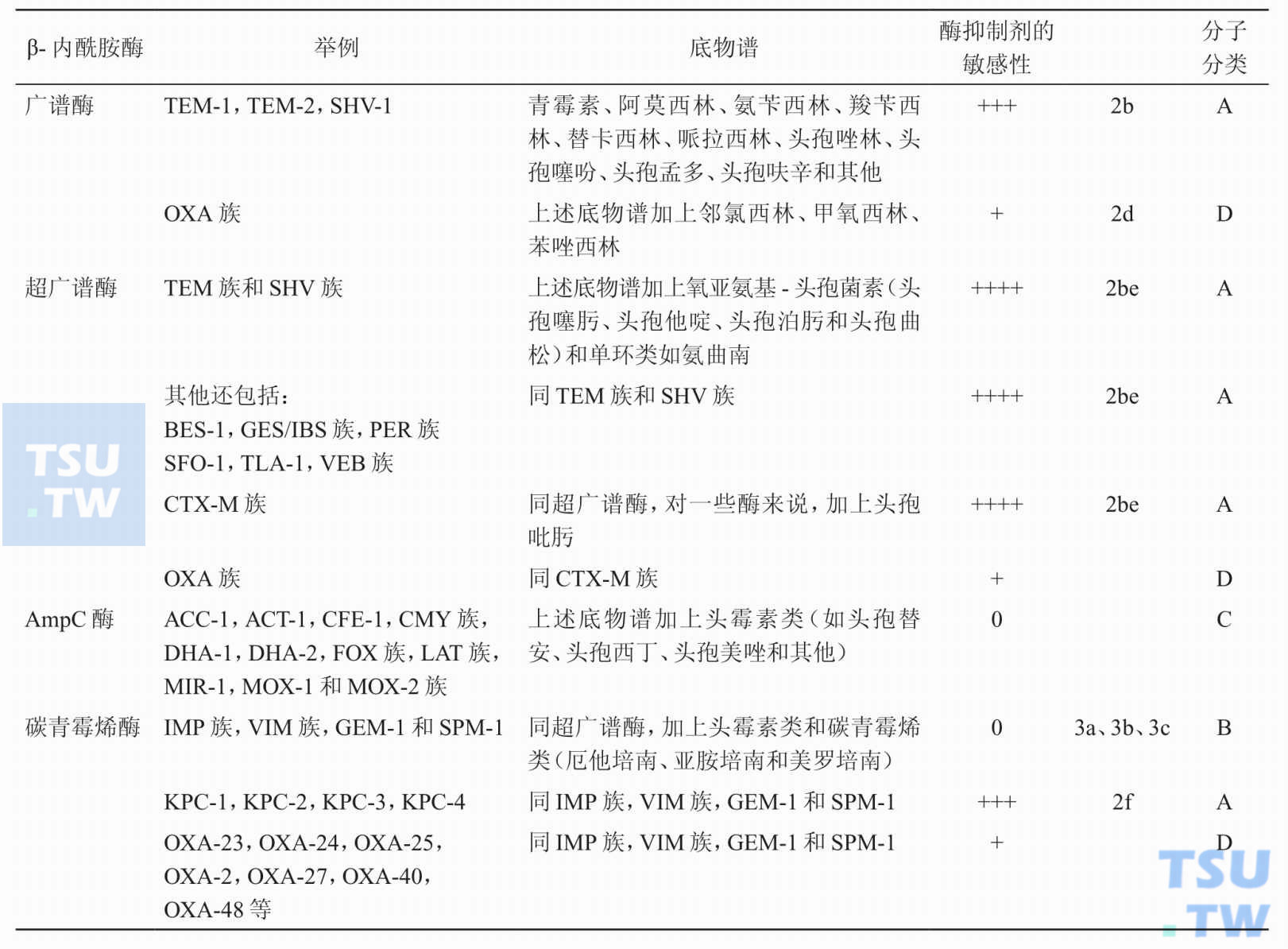
-
有关β-内酰胺酶的分类、底物谱以及对酶抑制剂的敏感性,了解了多少?
答:按Bush-Jacoby功能分类法可分为广谱β-内酰胺酶、超广谱β-内酰胺酶,AmpC酶和碳青霉烯酶。又可根据编码酶的基因、同源性等遗传学来源分成许多族、组和亚型。具体
1 -
KPC酶仅在肺炎克雷伯菌中发现吗?
答:目前该类酶已在肠杆菌科的多个菌属中有报道。除了肺炎克雷伯菌,还有阴沟肠杆菌、大肠埃希菌、产酸克雷伯菌、沙门菌属、弗劳地枸橼酸杆菌和铜绿假单胞菌等。该类酶除阴沟肠
2 -
目前已报道KPC酶有几种亚型?
答:有12种,包括KPC-1~KPC-12。它们之间只有个别氨基酸发生了突变。最初的KPC-1是美国于1996年在一株对碳青霉烯类耐药的肺炎克雷伯菌中发现一种能水解碳青霉烯类的丝氨酸酶,并称
3 -
什么是KPC酶?
答:KPC酶是指在肺炎克雷伯菌中产生的碳青霉烯酶,分子分类法(Amblar)的A组丝氨酸酶、功能分类法(Bush)的2f组,主要由质粒介导,其酶活性可受酶抑制剂抑制。产KPC型碳青霉烯酶可导致肺
4 -
D类碳青霉烯酶有哪些特征?
答:D类β-内酰胺酶,即OXA酶,它对苯唑西林的水解活性很强。2000年,来自苏格兰、西班牙、比利时、新加坡的亚胺培南耐药的鲍曼不动杆菌中,人们发现具有水解碳青霉烯类活性的D类
5 -
A类碳青霉烯酶有哪些特征?
答:A类碳青霉烯酶少见。它包括阴沟肠杆菌、黏质沙雷菌中由染色体介导的NMC-A、Sme-1~Sme-3、IMI-1酶,以及肺炎克雷伯菌、铜绿假单胞菌中质粒介导的KPC-1、GES-2酶。这类酶都是
6 -
什么是丝氨酸碳青霉烯酶?
答:目前已报道的丝氨酸碳青霉烯酶有两类。一类是属分子分类A组或功能分类2f组。属于A类酶的碳青霉烯酶主要存在于阴沟肠杆菌(NMC-A)、黏质沙雷菌(Sme-1)。另一类为近年来在鲍曼不
7 -
金属酶是否可由质粒介导?
答:金属酶大多是由染色体介导的,但不完全是。在20世纪90年代早期,日本首先报道了一株铜绿假单胞菌带有质粒介导的金属酶IMP-1,该酶并可通过转座子传播到沙雷菌和其他革兰阴性杆
8 -
目前已经报道的金属碳青霉烯酶包括哪些种类?
答:迄今网上公布的资料金属碳青霉烯酶IMP型酶23种和VIM型酶18种。前者根据DNA序列可分成两组:第一组IMP-1、IMP3~7、IMP9~11、IMP-16,第二组IMP-2和IMP-8、12、13、19、20;后者可
9 -
3b组的金属酶有哪些特征?
答:3b组金属酶特别易于水解碳青霉烯类抗生素,包括洋葱伯克霍德尔菌和芳香黄杆菌中发现的碳青霉烯酶以及气单胞菌中的AsbM1和CphA酶,这些酶除了活性部位的锌离子外,还需要更多锌
11 -
3a组的金属酶有哪些特征?
答:3a组金属酶可水解青霉素类、头孢菌素类和亚胺培南,该组酶包括蜡样芽孢杆菌酶、脆弱拟杆菌产生的CcrA、嗜麦芽窄食单胞菌产生的L1以及质粒介导的AMP-1酶,导致细菌对除单环&be
12 -
什么是金属碳青霉烯酶?
答:多数碳青霉烯酶为金属酶。该类酶属分子分类(Amblar)B组,因此亦称B类酶;或属功能分类(Bush、Jacoby)3组,为染色体介导,并可为EDTA所抑制,存在于多种不同革兰阴性细菌中。金属酶可水
13 -
质粒介导的AmpC酶和染色体介导的AmpC酶有什么关系?
答:CMY-2组成员与CMY-2的同源性达97.9%~99.7%,与CMY-2的差异为1~8个氨基酸,它们与染色体介导的弗劳地枸橼酸杆菌AmpC酶关系密切,其中CMY-2与染色体介导的弗劳地枸橼酸杆菌的AmpC酶
15 -
AmpC酶可以由质粒介导的吗?
答:可以。近年来已发现染色体上编码的AmpC酶基因与质粒结合并通过质粒传播到大肠埃希菌、肺炎克雷伯菌等肠杆菌科细菌中。根据氨基酸序列的同源性,这些酶可归为6个组。CMY-1组
16 -
染色体介导的AmpC酶一定是诱导酶吗?
答:不完全是。诱导性产AmpC酶革兰阴性杆菌,有较高频率(10−6~10−9)的自发突变。突变可发生在amp操纵子上:①ampC基因突变或ampC基因扩增;②ampC启动子和(或)衰减子突变;③
17 -
amp操纵子和AmpC酶的关系是什么?
答:amp操纵子是一个广为接受的β-内酰胺酶表达调节模型,它存在于许多革兰阴性杆菌如铜绿假单胞菌、肠杆菌属、枸橼酸杆菌属、沙雷菌属、普罗威登斯菌属及摩根菌属等的染色
18 -
什么是AmpC酶?
答:AmpC酶是染色体介导的β-内酰胺酶,由ampC基因编码,常见于肠杆菌属、枸橼酸菌属、沙雷菌属、铜绿假单胞菌属和蜂房哈夫尼亚菌。AmpC酶属于分子分类法的C组和功能分类法的
19 -
是否存在耐酶抑制剂SHV型β-内酰胺酶?
答:SHV-10是目前唯一的SHV型IRBLs,它来源于SHV-5,对抑制剂低水平耐药。此外,OHIO-1 型β-内酰胺酶也对酶抑制剂耐药,它与SHV-1的同源性达93%。
20