-
为什么细菌之间会传播耐药性?
答:细菌可通过质粒或转座子等方式,从其他细菌中获得外源性耐药基因,使自身遗传物质发生改变并获得耐药表型。这些外源性耐药基因可以通过染色体垂直传播,也可以以水平传播方式转
1 -
什么是细菌对抗菌药物产生耐药的生物化学机制
答:细菌耐药性的生物化学机制有以下6种:
2
① 产生抗生素水解酶:如β-内酰胺酶,细菌产生的β-内酰胺酶可与β-内酰胺环发生反应,导致抗菌药物被水解;
② 钝化酶产生:细 -
为什么细菌会在抗菌药物存在的环境中获得耐药性
答:细菌在分裂繁殖过程中会出现自发性基因突变,大部分基因突变对细菌本身并没有好处甚至有害,但有很小比例的突变可使细菌后天获得对某些抗菌药物的耐药性,使其在含有耐药性抗菌
3 -
为什么某些细菌会对抗菌药物存在天然耐药
答:天然耐药(natural drug-resistance)是某个菌种或菌属中所有菌株所表现出的内在特征,无论这些菌株来源于何处,他们对特定类型的抗菌药物均呈耐药。这种天然耐药性的产生可能是
4 -
为什么细菌以生物膜形式存在时耐药性会明显增强
答:生物膜(biofilm)是指细菌黏附于接触表面,分泌多糖基质、纤维蛋白、脂质蛋白等,将其自身包绕其中而形成的大量细菌聚集膜样物,是细菌为适应自然环境而产生的一种生命现象。生物
5 -
细菌耐药预警机制和干预措施具体应如何操作?
答:预警主要针对细菌耐药突出的抗菌药物而言,但每个抗菌药物又有比较宽的抗菌谱,在设定预警时必须注意对主要目标菌。一般来讲,预警可以分为两类:耐药信息预警和抗菌药物预警,耐药
6 -
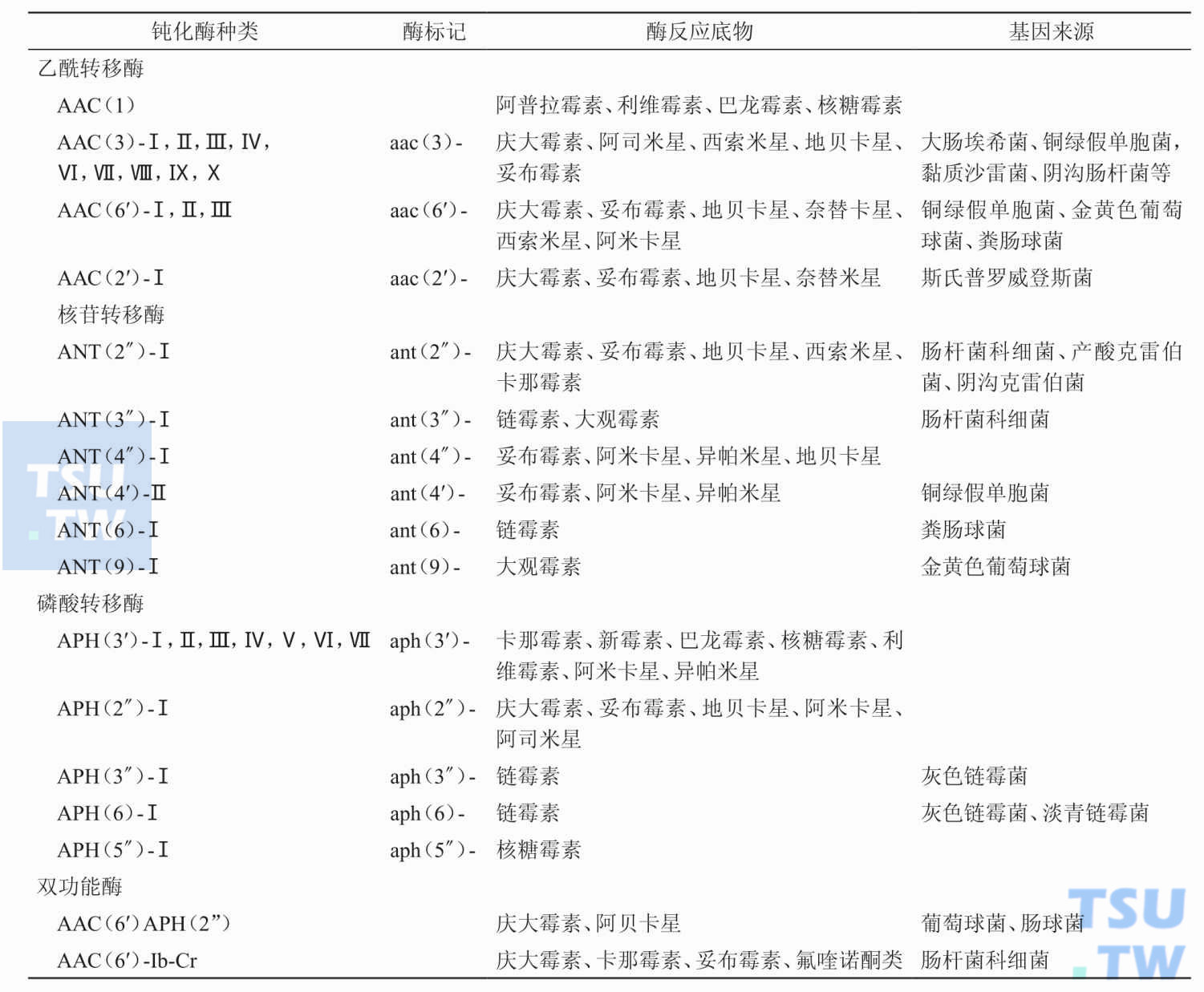
目前已知的钝化酶有哪几类?
答:目前已知有三类钝化酶,它们是:①乙酰转移酶(AAC);②磷酸转移酶(APH);③核苷转移酶(AAD或ANT)。三类酶又可按照所破坏的抗生素不同和作用点的不同而分为许多种。目前已知至少存在着30
7 -
氨基糖苷钝化酶所致的细菌耐药性
氨基糖苷类抗生素分子结构中都有2个或3个氨基糖分子和氨基环醇环,由配糖键相连接。细菌对氨基糖苷类抗菌药物的耐药机制非常复杂,主要包括细胞内外电位差的改变、细菌的主动外
8 -
是否存在耐酶抑制剂SHV型β-内酰胺酶?
答:SHV-10是目前唯一的SHV型IRBLs,它来源于SHV-5,对抑制剂低水平耐药。此外,OHIO-1 型β-内酰胺酶也对酶抑制剂耐药,它与SHV-1的同源性达93%。
9 -
为什么耐酶抑制剂β-内酰胺酶又称耐酶抑制剂TEM型酶(IRT)?
答:核苷酸序列分析表明这些酶从TEM-1或 TEM-2演化而来,故亦曾称为耐酶抑制剂TEM系列酶(inhibitor-resistant TEM,IRT),按TEM系列命名。此类酶属于分子分类法的A类和功能分类法的2b
11 -
什么是耐酶抑制剂β-内酰胺酶(IRBLs)?
答:一种耐酶抑制剂克拉维酸的β-内酰胺酶。最先是1989年在法国、德国的大肠埃希菌中发现的,现在肺炎克雷伯菌、奇异变形杆菌、弗劳地枸橼酸杆菌和产酸克雷伯菌中也有发现
12 -
什么是SHV型β-内酰胺酶?
:SHV是“去巯基”的英文(sulfhydryl variable)缩写,SHV型酶可以水解β-内酰胺类抗生素的巯基。临床分离的肺炎克雷伯菌中,90%的菌株可以产生染色体介导的SHV-1&bet
13 -
什么是D组β-内酰胺酶?
答:即Ambler分类法的D组β-内酰胺酶,即苯唑西林水解酶,此酶为染色体和质粒介导皆有。主要水解氯唑西林和苯唑西林。该类酶在Bush分类法中属2组中的2d组。
14 -
什么是C组β-内酰胺酶?
答:即Ambler分类法的C组β-内酰胺酶,如AmpC酶,亦属Bush分类法的1组酶,染色体介导,相对分子量约39 000,主要水解头孢菌素类。在其活性部位亦带有丝氨酸,但与A组酶缺乏同源序列。
15 -
什么是B组β-内酰胺酶?
答:即Ambler分类法的B组β-内酰胺酶,如金属酶,亦属Bush分类法的3组酶,染色体介导,相对分子量为56 000,其活性部分是结合锌离子的硫醇基。主要水解碳青霉烯类及β-内酰胺类
16 -
什么是A组β-内酰胺酶?
答:即Ambler分类法的A组β-内酰胺酶,亦属Bush分类法的2组酶,质粒介导、相对分子质量约29 000,其活性部位具有一丝氨酸残基,主要水解青霉素类,可为β-内酰胺酶抑制剂所抑制
17 -
目前有关β-内酰胺酶分类的系统有哪些?
答:以Ambler根据氨基酸组成和核苷酸序列的分子分类法和Bush、Jacoby与Medeiros(1995)根据酶的底物谱和克拉维酸的抑酶作用等特点进行的功能分类法最为广泛采用。前者将β-
18 -
β-内酰胺酶所致的细菌耐药性
细菌对β-内酰胺类抗生素产生耐药性大凡可由下列机制所致。青霉素结合蛋白(PBPs)的改变,在革兰阴性杆菌中的外膜蛋白结构的改变和内膜上依赖能量的外排泵的产生,阻止抗菌药
19 -
什么是替代性靶位蛋白?与细菌耐药性有何关系?
答:细菌为抵抗抗菌药物的抑制作用而产生的一种与抗菌药物亲和力低的新的靶位蛋白,取代正常的靶位蛋白与抗菌药物结合。如甲氧西林耐药的金黄色葡萄球菌由mecA基因编码产生一种
20