-
精子运动能力的发育控制:精子细胞信使系统
附睾精子成熟过程中,与精子运动能力获得和发育调控关系比较密切的精子细胞信使系统,目前认为有钙离子、钙调蛋白、cAMP、磷酸二酯酶抑制剂以及细胞外低浓度ATP的非能量调节作
181 -
精子游动的供能中心:附睾精子的能量系统
精子中大量的线粒体构成精子尾部中段螺旋形排列的线粒体鞘,是精子的供能中心。精子糖酵解和线粒体呼吸代谢中产生的ATP是精子运动的主要能量来源,能激活精子的鞭毛系统。在大
182 -
精子能够运动的结构因素
与附睾精子运动能力获得的相关因素很多,也很复杂。目前概括起来可以归结为四个方面:①精子附睾成熟运行过程中的结构调控,如精子鞭毛的结构包括巯基的变化;②附睾精子的能量系统
183 -
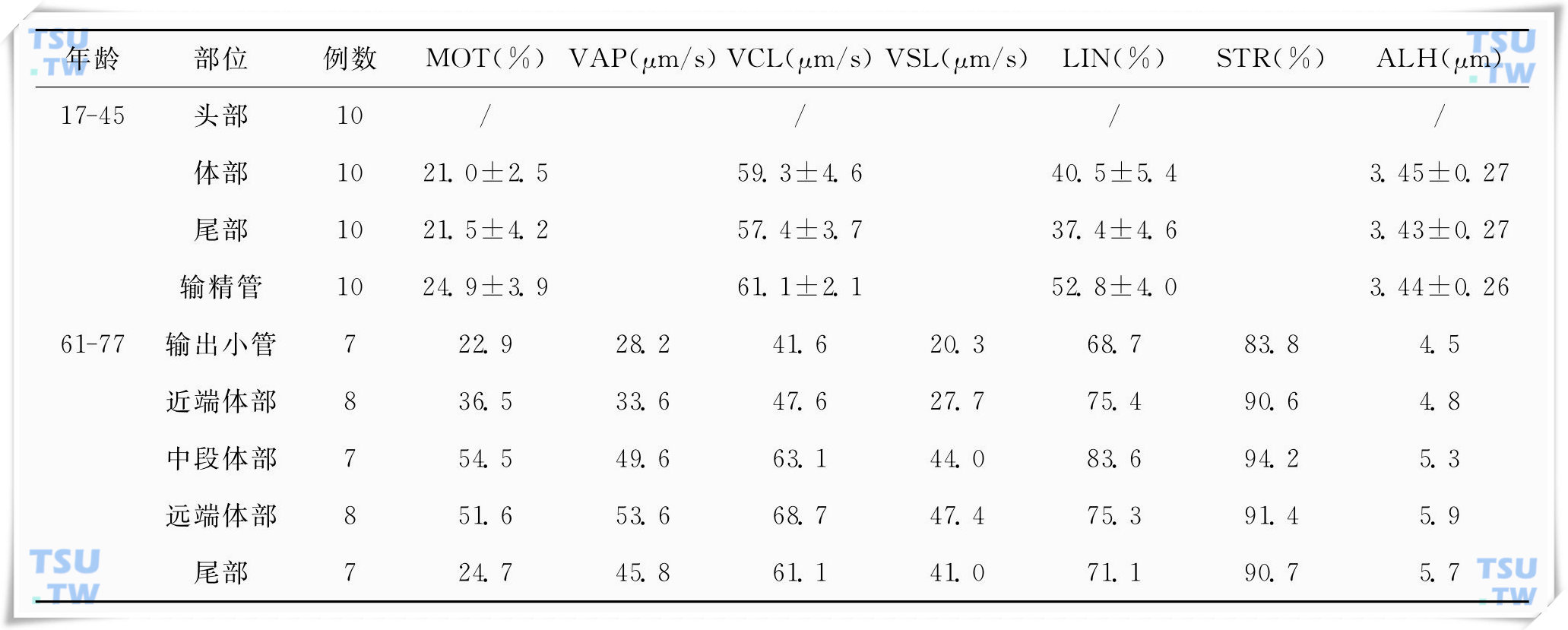
附睾精子成熟过程中运动能力获得的表现
精子进入雌性阴道并快速通过子宫颈进入子宫,最后到达输卵管。目前已比较明确,这种精子在女性生殖道内的运行,除了受女性生殖道的蠕动或收缩所给予的推动力之外,主要是依靠精子自
184 -
附睾精子成熟过程中的能量代谢
精子活动和代谢的客观测量显示活动是由糖酵解和线粒体呼吸作用产生的ATP所支持的。附睾精子含有所有能有效进行糖酵解的酶类,其中大多数是处于一个高活性状态。在大鼠附睾精
185 -
附睾精子成熟过程中巯基的变化
精子膜在附睾精子成熟过程中,膜上的巯基逐渐被氧化为二硫键。由于二硫键的键能很大(约125~418kJ/ml),他可把不同肽链或同一肽链的不同部分连接起来,对稳定膜蛋白质起了重要的作用
186 -
精子成熟过程中精子膜蛋白质组成的改变
在附睾精子成熟过程中,精子膜蛋白质组成发生了重大的改变,这种改变或是在运行过程中加进了新的蛋白质成分,用高离子浓度培养液洗涤大鼠和仓鼠成熟精子所得到表面蛋白比未成熟精
187 -
精子成熟:精子膜膜脂成分及代谢、膜脂流动性的改变
有关直接研究精子在附睾成熟过程中膜流动性的报道比较少。但现有结果一般认为是随附睾成熟运行,精子膜的流动性是逐步降低的。Majumder用芘(pyrene)掺入膜的脂相形成在480nm的
188 -
精子成熟过程中精子膜表面糖基和表面负电荷的改变
研究精子膜糖基的组成,一般可用植物凝集素(lectin)作为细胞膜的特异性分子探针。如刀豆球蛋白(ConA)可与葡萄糖胺和甘露糖结合,麦芽凝集素(WGA)可与N-乙酰葡萄糖胺结合,蓖麻凝集素(RCA
189 -
精子成熟过程中精子膜通透性的改变
精子在附睾成熟过程中精子膜的结构也发生了重要的变化,膜上成分以及生化、生理、物理性状的明显改变。包括膜的通透性、膜的流动性、膜电荷、膜脂、膜蛋白以及膜上糖基成分和
190 -
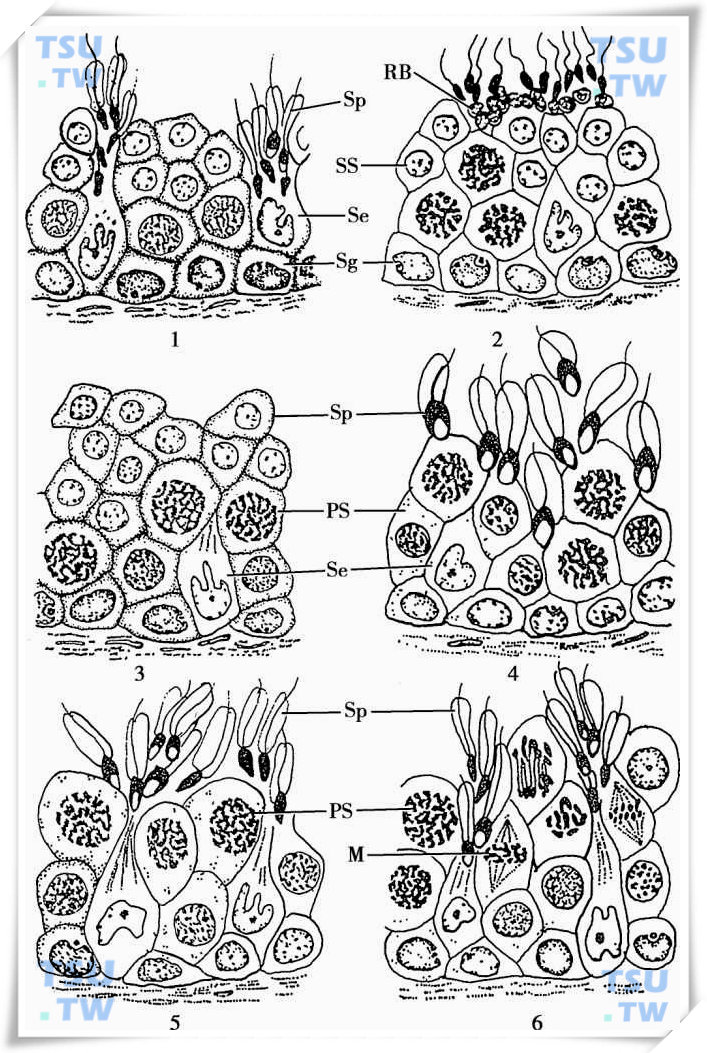
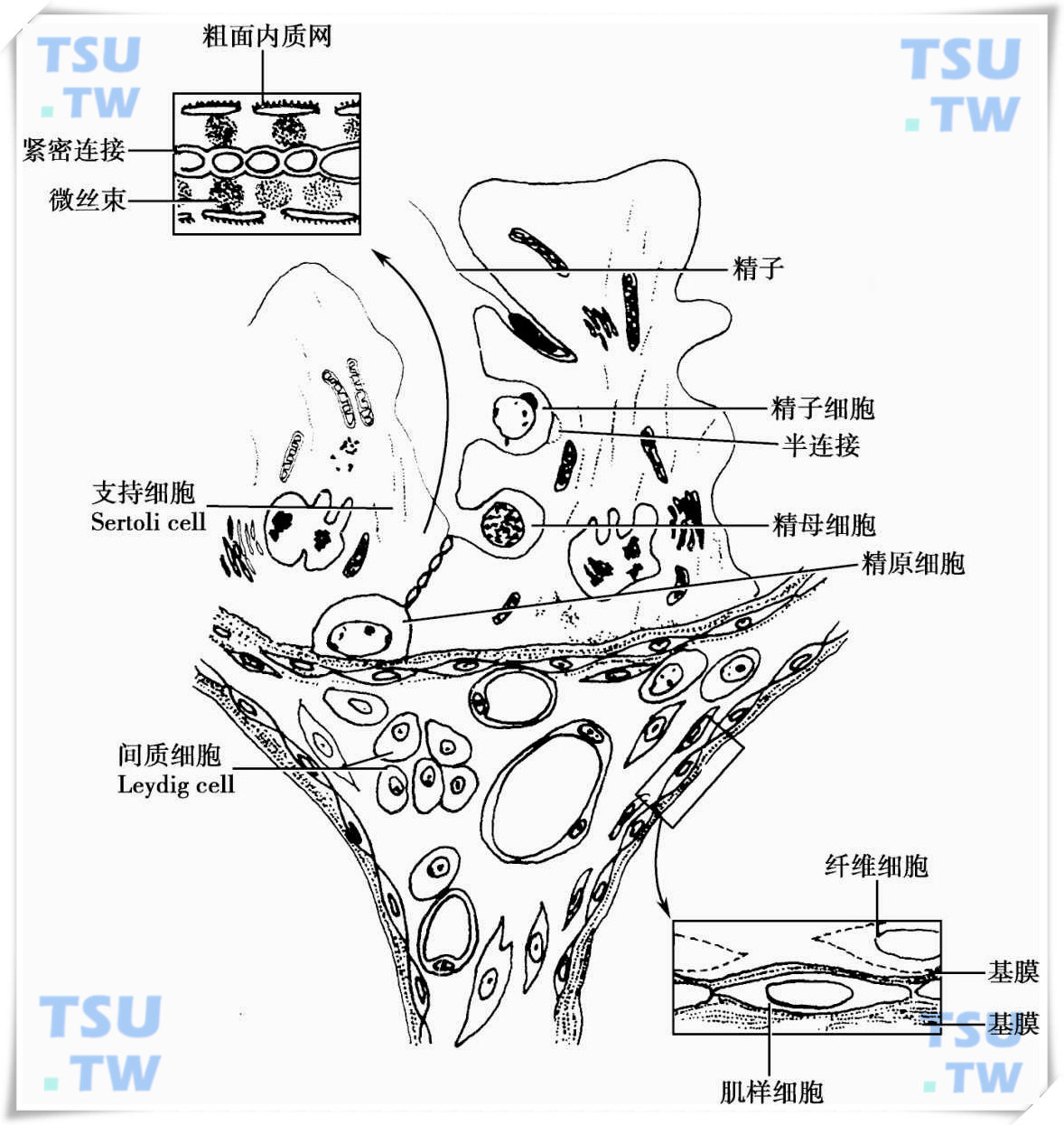
精子发生的动力学:效率、时程、规律、生精上皮周期
精子发生的效率生精上皮由Sertoli细胞和生精细胞组成,在灵长类和大鼠的生精上皮中,其Sertoli细胞和生精细胞的数量明显不同。大鼠Sertoli细胞占10%~20%,生精细胞占50%~80%;在人类
194 -
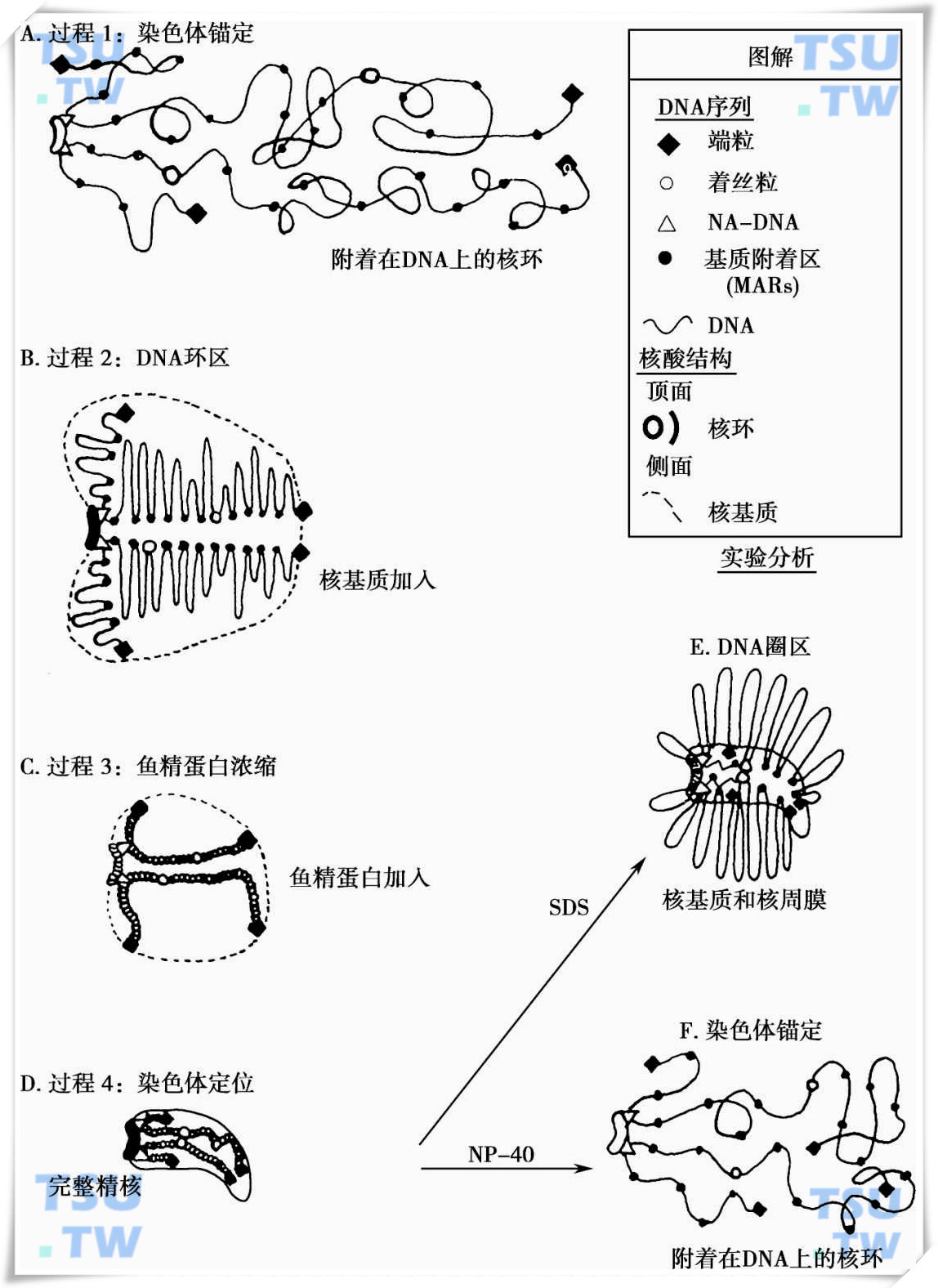
精子形成的染色体结构
以往一直以为,在精子形成后,精子的染色体组被压缩成如同晶体般紧缩的结构并失去其活性,只有在受精解螺旋后才具有活性。随着近几年来关于精子DNA结构和功能研究的发现,提示精子
195 -
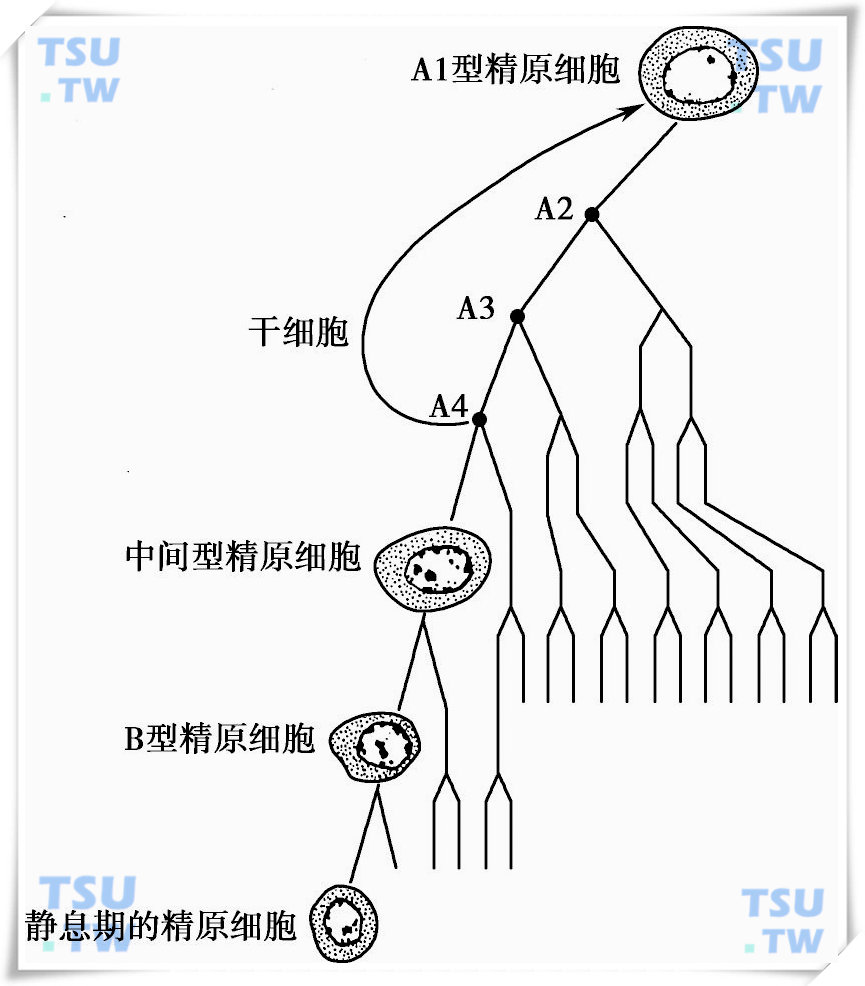
精子发生的有丝分裂增殖的过程
从精原干细胞(spermatogonia type A stem cell)形成高度分化和特异的精子是一个极其复杂的细胞分化过程,由精原干细胞的增殖分化、精母细胞的减数分裂和精子形成3个阶段组成。
197 -
精子膜的离子通道
人精子的成熟、运动、获能和精-卵结合是一个复杂的过程,很多因素影响到这些功能,其中多种离子起到重要作用,如Na+、K+、Ca2+、Cl-等,这些离子又大多通过精子细胞膜上的离子通道
198 -
精子膜的脂质成分
人精子胞膜的脂质组成人精子胞膜的磷脂种类、组成与体细胞基本一致,磷脂约占70%,其余为中性脂质(25%)和糖脂(5%)。磷脂又分为磷酸甘油酯(phosphoglycerolipids)和鞘磷脂(sphingomyeli
200