
在精子发生的过程中,最明显的形态学变化是精子细胞的改变。包括:高尔基复合体囊泡融合成一个大的顶体囊泡,覆盖于精子细胞核大部分的表面,形成顶体;中心粒迁移到细胞核的尾侧,远端中心粒分化形成精子尾部的鞭毛中轴;线粒体集中在鞭毛起始部,形成线粒体鞘;多余的胞质形成残余体而弃之,如下图所示。
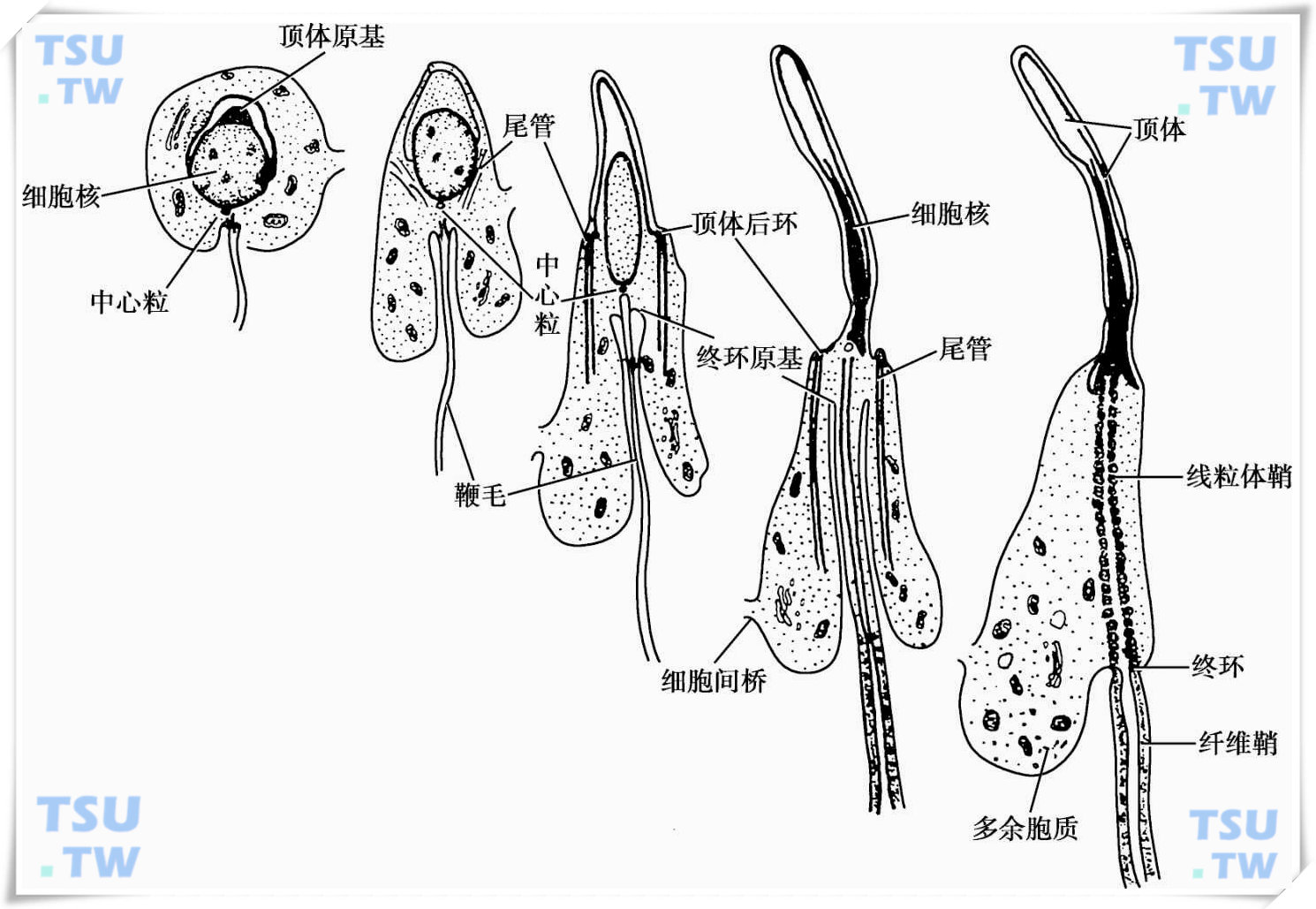
豚鼠精子细胞变态过程示意图
在精子形成早期,胞质内含有大量的高尔基复合体。随着精子形成(spermiogenesis)过程的开始,高尔基复合体产生圆形囊泡,称之为前顶体囊泡,囊泡内含有致密的颗粒,称之前顶体颗粒。随后前顶体囊泡逐渐融合形成了一个大的顶体囊泡。随着精子细胞核的浓缩变长,顶体囊泡成为扁平状,覆盖在细胞核的表面,由顶部向尾部逐渐包绕精子细胞核的大半部,形成顶体。
在精子细胞形态变化中,较为明显的是细胞能量供应器——线粒体的改变。在精子形成刚刚开始的时候,精子细胞中的线粒体形态已完全不同于精原细胞或体细胞的线粒体形态,此时的线粒体嵴贴附在线粒体膜的周围,其间为弥散的和空泡化的基质。当线粒体组成精子尾部线粒体鞘时,线粒体的形态为月牙状。随着线粒体结构的改变,线粒体的蛋白成分也随之改变。在精子形成完成时,精子细胞多余的胞质已被除去,在留下的很少的胞质中不含核糖体,胞质蛋白质的合成停止了,但线粒体蛋白质的合成仍在继续。形态统计显示,在精母细胞和精子细胞中线粒体数超过103个/细胞,但每个精子中段仅容纳约75个线粒体,表明在精子形成的过程中大量的线粒体被遗弃。
在细胞核浓缩时,原先与DNA结合的组蛋白被高碱性的过渡蛋白替代,进一步又被精核蛋白替代,精核蛋白能中和DNA电荷,降低DNA分子间的静电排斥作用,并通过二硫键的交联形成致密的细胞核。
在精子发生过程中,生精细胞的核染色体组成及结构在精原细胞向精母细胞及精子细胞的发育过程中发生了极大的变化,其间以核小体组蛋白为基础的结构逐步被排列紧密的精核蛋白代替,通过一系列的变化,形成新的染色质重组。在雄鼠干细胞的减数分裂前组蛋白的合成有两个波峰,分别在细线前期和粗线期。鼠睾丸精原细胞表达睾丸特异的H3蛋白,细线前期表达tH2A和tH2B蛋白,粗线期表达H1t蛋白。减数分裂后的精子细胞特异表达H2B蛋白。精子细胞变形过程中,通过蛋白质转化,使精核蛋白替代核小体上的组蛋白。在鼠和人类,转化蛋白有TP1和TP2(transition proteins 1and 2)两种,精核蛋白有P1和P2(protamines 1and 2)两种。
精核蛋白与DNA之间的联系导致染色质形成层层折叠的结构,使精子细胞核高度浓缩,其DNA至少比有丝分裂染色体多六次折叠。在精子细胞的基因组发生浓缩时,其转录活性消失,但仍有少量蛋白质的合成,精子细胞中mRNA的贮存以便用于翻译。组蛋白向精核蛋白转化过程的异常可导致不孕或胚胎早期死亡。
整个精子发生的过程是在Sertoli细胞围成的微环境中进行的,发生中的各级生精细胞嵌入在Sertoli细胞质的深处并与Sertoli细胞膜形成独特的半连接接触(下图)。在精子发生过程中,生精细胞在Sertoli细胞作用下,由生精小管的基底小室通过Sertoli细胞之间的紧密连接移向近腔小室。当精子细胞完成变形后,被Sertoli细胞释放到管腔,浸泡在睾丸液中,并随睾丸液的流动,通过睾丸的输出小管,储存在附睾中。
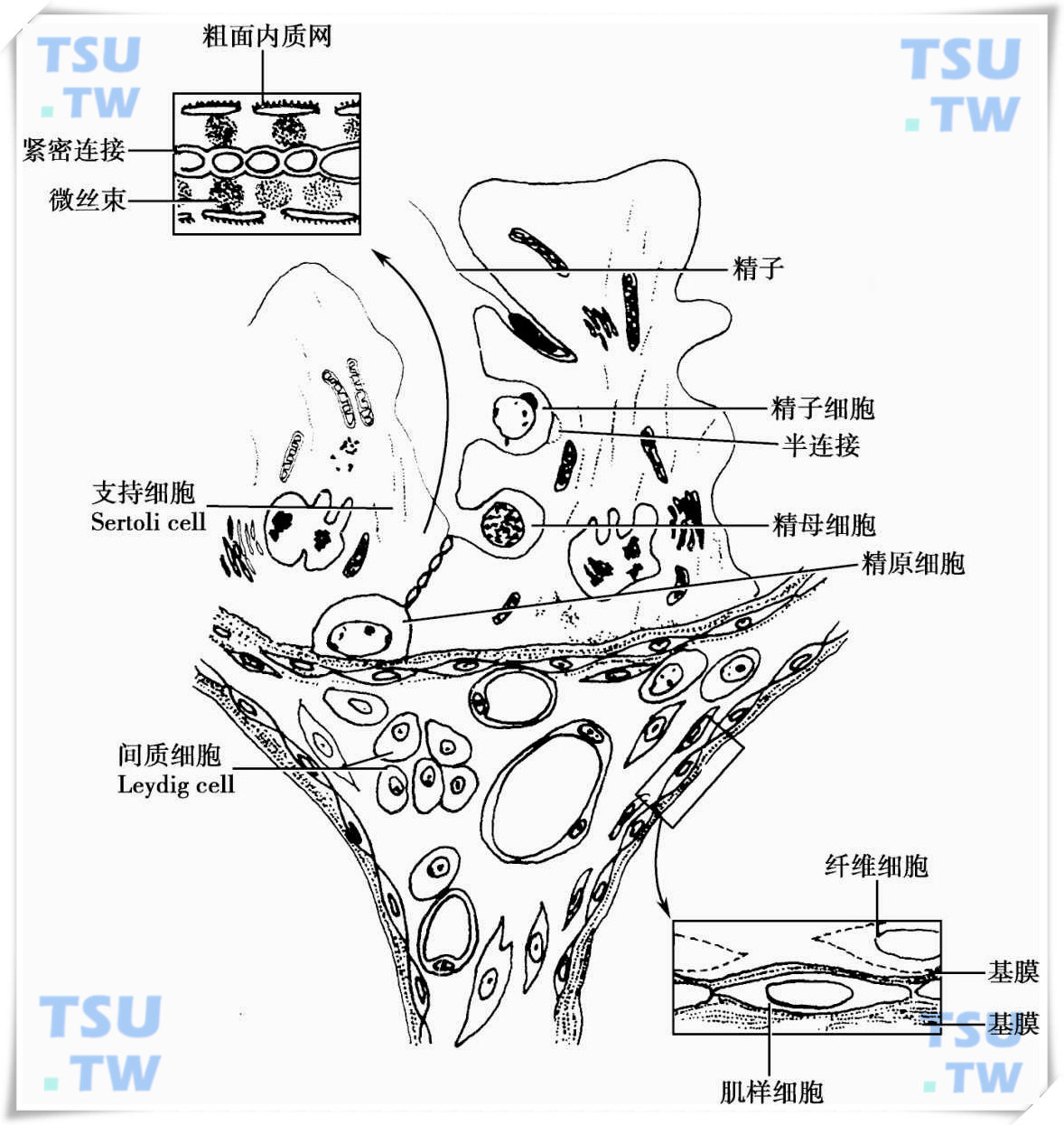
成年睾丸横切面模式图
每一个精子都是一个大的克隆的成员之一,这个群体是由一个A型精原细胞通过多次的有丝分裂增殖和减数分裂而来的,但在这个群体中,没有完全相同的精子。第一次减数分裂时,初级精母细胞中来自于父方与来自于母方的23对染色体之间按223随机组合被分到次级精母细胞中,就会形成223=8 388 608种不同的染色体组成。此外,在每一条染色体上都有许多基因,其中许多基因都相互连锁,由于联会时,同源染色体之间可能发生染色单体的部分片段交换,产生新的连锁关系。减数分裂时非同源染色体之间的随机组合,同源染色体之间的相互交换,确保了每一个精子具有自己独特的遗传信息,这是人类表现出复杂的遗传和变异现象的基础。
生精细胞分化产生功能成熟精子的过程很长,在此期间,许多分化步骤常常由于各种因素干扰而发生改变,造成精子发生过程中的遗传损伤和结构异常。在减数分裂中,尽管具有一系列复杂的保护措施,包括DNA配对、DNA修复等,但时常还会发生易位和非整倍体。统计资料表明在正常人的精子中,1号染色体双体发生率为0.09%,2号为0.08%,4号为0.11%,9号为0.14%,12号为0.16%,15号、16号和18号为0.11%,20号为0.12%,21号为0.29%,性染色体为0.43%。不育症患者精子中,除总的染色体双体数目增多外,性染色体双体的数目显著升高。因此,不育男性中有相当一部分病人是由于生精细胞分裂阻滞在减数分裂期,这些病人几乎绝大多数为染色体异常配对和等位基因分离。在精子形成的过程中,对于正在成熟的生精细胞也是极易产生遗传和结构缺陷的。在精子形成时,DNA的修复能力已降低,此时组蛋白从核小体上被替换下来,能导致单倍体基因组DNA的损伤;此外,由于大量的形态学改变,均可能使生精细胞阻滞在精子形成阶段而导致男性不育,如精子的轴丝缺少、中部的线粒体异常或尾部的装配异常均可导致精子运动障碍;与精子核形成相关的蛋白质突变将导致精子头部畸形。
生精小管界膜由基膜、肌样细胞和成纤维细胞等部分构成。肌样细胞间有点状的细胞连接,没有血管、淋巴管和神经穿过界膜进入生精小管。生精小管中Sertoli细胞间的紧密连接复合体分隔近腔小室和基底小室。在紧密连接两侧胞质中存在内质网池,称表面下池,与粗面内质网相连续;在表面下池和紧密连接之间存在着相互平行的微丝束,这是Sertoli细胞连接中的收缩系统,可开启Sertoli细胞的紧密连接,使正在发育的生精细胞从基底小室移向近腔小室;精母细胞、精子细胞和精子位于近腔小室,并与Sertoli细胞存在密切的联系,形成半连接。
