
男性胚胎第7周时,在睾丸决定因子(testis-determining factor,TDF)的影响下,未分化性腺开始性别分化,其皮质退化消失,而髓质增殖发育,在此基础上,睾丸形态逐渐构建,各种细胞分化形成。
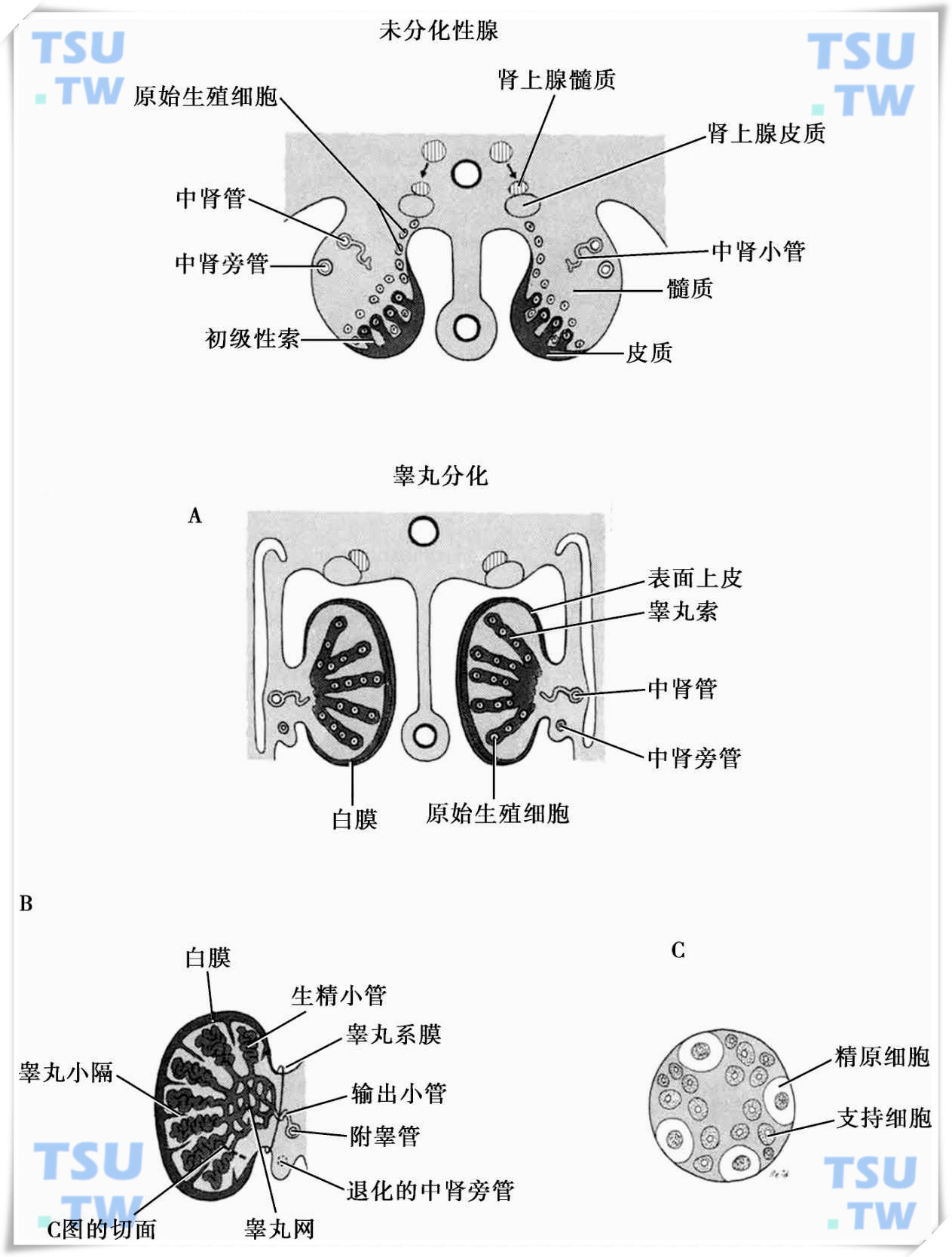
睾丸发生与分化模式图
白膜与血管膜
男性胚胎第8周时,表面上皮与髓质之间的间充质分化为一层较厚的致密结缔组织,即白膜(tunica albuginea),它的出现是胎儿睾丸发生的一个重要标志(图A)。人胚第12~13周,在睾丸的上皮深面,白膜已可见数层纤维;14周后,白膜内出现PAS阳性纤维,排列较致密。血管膜于14周出现于白膜深面,随后血管膜中出现较大的血管,血管膜和白膜的厚度均逐龄增加。
生精小管形成及其细胞分化发育
在TDF的作用下,髓质中的初级性索不断生长,与表面上皮分离,向髓质深部增殖发育为睾丸索(testicular cord)(图A)。睾丸索以后发育为生精小管和直精小管,其末端断裂吻合成睾丸网。在人类,胚胎第4个月时睾丸索即已分化形成细长、弯曲的袢状生精小管(图B),但从胎儿期直至出生后青春期之前,生精小管始终是没有管腔的实心细胞索。
目前对于人类原始生殖细胞分化发育的时间和过程研究较少,有报道显示,随着生精小管的形成,早期散在于睾丸索中央或周边的原始生殖细胞逐渐移位于管壁上皮细胞基底部,此时其胞质内仍含显著而多少不等的糖原颗粒,并且碱性磷酸酶反应阳性;24周后,沿基膜分布的少数原始生殖细胞,核着色深,显示出精原细胞的特征,可称为精原细胞(spermatogonia)。对小鼠原始生殖细胞分化的研究相对较多。报道显示,胚胎第13.5d的雄鼠性腺已经开始分化,可以见到实心的原始生精小管结构;胚胎第16d时,可观察到生精小管细胞索中含有两种细胞,原始生殖细胞(PGCs)位于索中央,支持细胞(Sertoli cell)位于索周边贴于基膜上;出生后第1d的新生小鼠,可见大多数PGCs仍在索中央,但部分PGCs迁向基膜,有学者认为此时的生殖细胞可称为生殖母细胞(gonocyte);至出生后第6d,生殖细胞绝大部分迁移至基膜,分化形成原始的A型精原细胞(primitive type A spermatogonia);出生后第8d,生精上皮中可见到A型精原细胞(type A spermatogonia)和B型精原细胞(type B spermatogonia);此后直至青春期时,精原细胞才分化形成精母细胞,经过减数分裂和分化变形,最终生成精子。
在PGCs发育分化的同时,来源于初级性索的支持细胞也逐渐分化形成。此时支持细胞排列无一定方向,胞体小而边界不清,核长圆形着色深(图C)。在胎儿睾丸中,支持细胞优先占据了生精上皮的大部分。但从人胚20周至32周,支持细胞与生殖细胞之比约从3∶1减少为2∶1。至新生儿期,仅有少数支持细胞,此时细胞核呈三角形,着色深,胞质内可见少许张力原纤维,但其仍不具备性成熟期支持细胞的典型结构。此后直至青春期前,有关人支持细胞的形态结构变化很少见报道。而在小鼠中,有研究者观察了胚胎期和青春期前不同阶段的睾丸中支持细胞的分化发育,结果显示:胚胎第13d,可见支持细胞分散于PGCs之间,呈不规则形状,核圆形或卵圆形,染色浅;胚胎第16d时,支持细胞位于生精小管细胞索周边贴于基膜,核形态变为不规则,可见1至数个核仁;胚胎第18d时,细胞开始向索中延伸;出生后第1d,支持细胞多为长椭圆形,核中核仁明显,核膜可见有异染色质附着,细胞进一步向索中央突出;出生后第10d,支持细胞胞体向索中明显突出,核色浅不规则,明显可见核仁;大约在出生后第15d青春期开始时,生精小管管腔开始形成,支持细胞呈现成熟期的典型结构。
从胚胎期直至出生后青春期之前,睾丸中生殖细胞的有丝分裂和减数分裂都受到抑制,直到青春期时,才开始完成精子形成的所有过程,包括生殖细胞的有丝分裂、精原细胞的分化以及精子发生。现在认为,早期睾丸支持细胞和原始生殖细胞之间的相互联系可能起着重要作用,这两种细胞之间的相互作用,直接抑制了原始生殖细胞的有丝分裂,同时阻止生殖细胞进入减数分裂。此外,McLaren等在小鼠中的研究显示,睾丸索结构的形成是减数分裂抑制所必须的,不能形成睾丸索将导致生殖细胞进入减数分裂。
睾丸间质和间质细胞
分散在生精小管之间的间充质分化为睾丸间质及睾丸间质细胞(Leydig cell)。谷华运等通过观察6.5~8周的男性胎儿,发现在睾丸索和生精小管之间,除有丰富的血管和淋巴管等外,还可见到许多胞体较大、呈圆形或多边形、胞质呈嗜酸性的间质细胞。目前一致认为,间质细胞在人胚第8周时已出现,并分泌雄激素。此时的间质细胞为分化前型,主要特征是胞质中滑面内质网和线粒体开始增加,而粗面内质网减少。胚胎第9~14周时,睾丸间质细胞转化为分化型。胚胎第15~18周时,间质细胞发育为成熟型,其数量明显增加,占睾丸体积一半以上,并具有旺盛的合成分泌雄激素能力,形成了胚胎时期雄激素的分泌高峰。随后,间质细胞变性退化。人胚胎间质细胞变性速度缓慢,要低于猴和啮齿类动物,一般在出生后几周,睾丸内几乎见不到间质细胞。直到青春期,间质细胞重新增多增大,再次分泌雄激素。在睾丸发育的早期阶段,雄激素的合成受到胎盘分泌的人绒毛膜促性腺激素(human chorionic gonadotropin,hCG)的调节,hCG在胚胎第8~12周时出现高峰,刺激间质细胞合成分泌雄激素。睾丸发育到晚期阶段,垂体分泌的促性腺激素逐渐替代胎盘的作用。
