
减数分裂(meiosis)起始于细线前期的初级精母细胞,这一时期的男性生殖细胞进行最后的半保留DNA复制,在这些男性生殖细胞中所有的DNA合成均是DNA修复合成,作为减数分裂的早前期,染色体呈细线状,仅性染色体形成致密的异固缩小体。随着细线期(leptotene)和偶线期(zygotene)精母细胞从生精小管的基底小室通过Sertoli细胞的紧密连接进入近腔小室,初级精母细胞的核形态发生明显的变化。在细线期,虽DNA已复制,但染色体看不到两条姐妹染色单体,染色体丝折叠成珠状,形成染色粒(chromomere)。随着染色体的进一步折叠,染色粒越来越大,数目也越来越少,减数分裂进入偶线期。此时同源染色体配对,称为染色体联会(synapses)。染色体配对是高度特异的,在点与点、染色粒与染色粒之间配对是绝对准确和特异的,配对的同源染色体不互相融合,其间有0.15~0.32nm的间隙,间隙中主要成分是蛋白质,形成联会复合体。
联会复合体除帮助稳定同源染色体配对,更重要的是帮助同源染色体小片段之间的交换和重组。联会复合体是减数分裂细胞中一个高度保守的结构,自然界存在的生命万物中,无论是水中霉菌,还是人类都需要有效的联会,这是父源和母源染色体基因互换和重组的一个特异结构。在粗线期(pachytene)的精母细胞,随着染色体的进一步折叠,每条染色体出现两条染色单体,每条染色单体均有自己的着丝点。此时,在同源染色单体之间能发生某些片段的断裂和交换(crossing over),这一现象能在光镜下看到,称为交叉(chiasma)现象,也是染色体基因互换和重组的基础。在双线期(diplotene)时,同源染色体之间开始分离,但在交叉的部位仍连在一起。在终变期(diakinesis),核仁消失,交叉数目明显减少,同源染色体只有端部在一起。由终变期进入减数分裂的中期、后期和末期,每一个初级精母细胞分成为两个次级精母细胞。每一个细胞中容纳单倍体染色体数,每一个染色体由两条染色单体组成,两条染色单体靠着丝点处相连。
第一次减数分裂完成后,经过短暂的间期(有时此期并不存在),染色体不再进行复制即进入第二次减数分裂。在第二次减数分裂时,两条染色单体在着丝点处分开,各自移向细胞两端,在较短的时间里,次级精母细胞即分裂成为早期的精子细胞。
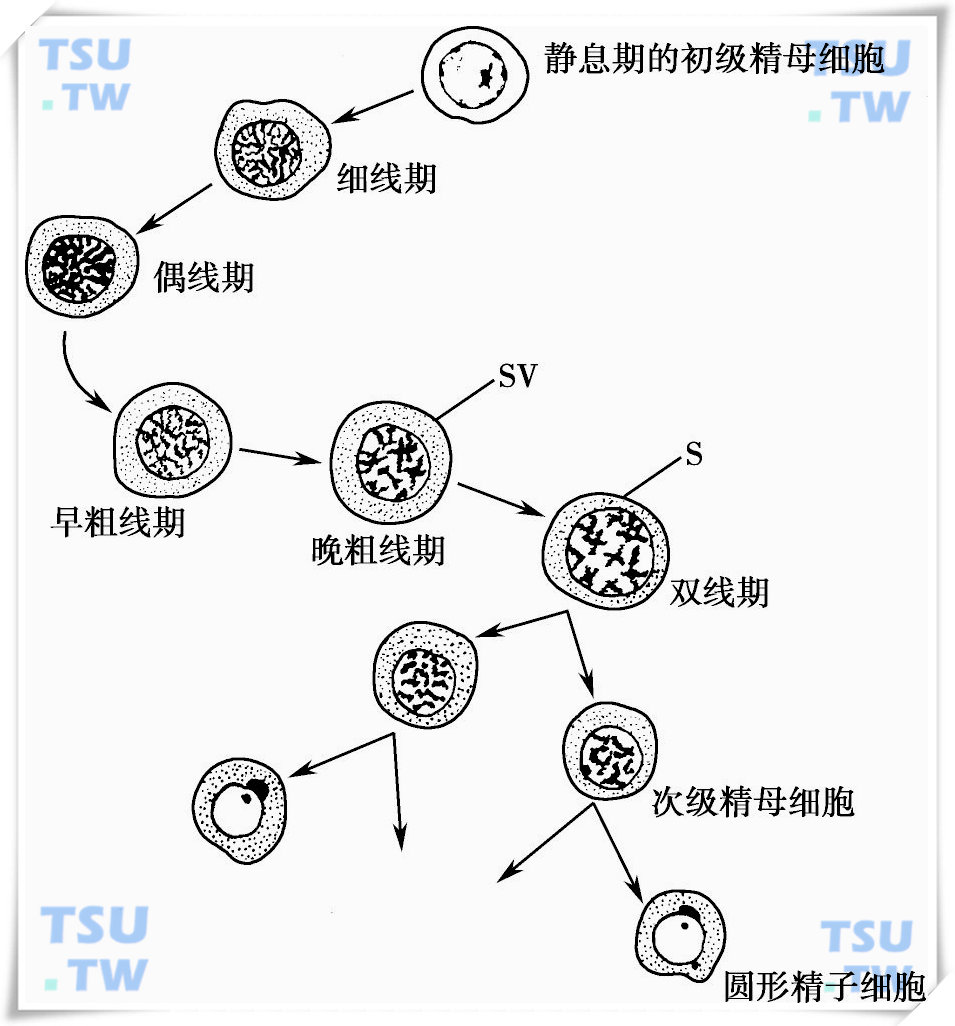
在近腔小室中静息期初级精母细胞减数分裂的过程
SV:指粗线期时,性染色体发生联会形成的性泡(sex vesicle);S:联会(synapses)
减数分裂是一个极其复杂的过程,需要许多新的结构蛋白和酶的参与,它们对染色体的排列、断裂、重组和修复等具有重要的意义。如:Rad51是与人类同源的作用于基因重组的细菌重组蛋白;ATM相关基因是预防DNA损伤的基因家族的一员;泛素偶联修复酶(ubiquitin conjugating repair enzyme)参与蛋白质转化;另外还有减数分裂特异性的DNA双链断裂酶、DNA重组蛋白及减数分裂特异的热休克蛋白等。
