-
目前已报道KPC酶有几种亚型?
答:有12种,包括KPC-1~KPC-12。它们之间只有个别氨基酸发生了突变。最初的KPC-1是美国于1996年在一株对碳青霉烯类耐药的肺炎克雷伯菌中发现一种能水解碳青霉烯类的丝氨酸酶,并称
151 -
KPC酶仅在肺炎克雷伯菌中发现吗?
答:目前该类酶已在肠杆菌科的多个菌属中有报道。除了肺炎克雷伯菌,还有阴沟肠杆菌、大肠埃希菌、产酸克雷伯菌、沙门菌属、弗劳地枸橼酸杆菌和铜绿假单胞菌等。该类酶除阴沟肠
152 -
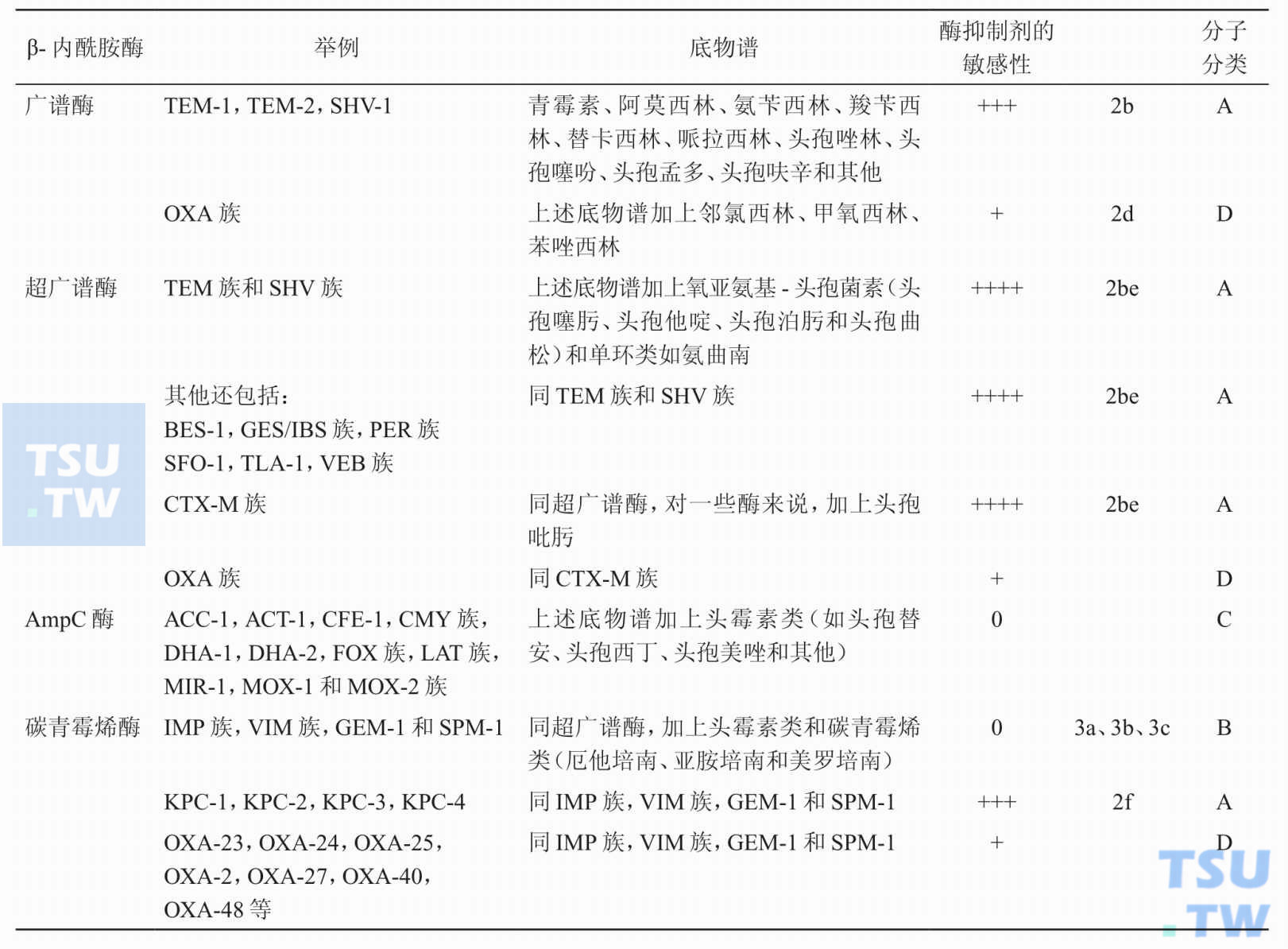
有关β-内酰胺酶的分类、底物谱以及对酶抑制剂的敏感性,了解了多少?
答:按Bush-Jacoby功能分类法可分为广谱β-内酰胺酶、超广谱β-内酰胺酶,AmpC酶和碳青霉烯酶。又可根据编码酶的基因、同源性等遗传学来源分成许多族、组和亚型。具体
153 -
氨基糖苷钝化酶所致的细菌耐药性
氨基糖苷类抗生素分子结构中都有2个或3个氨基糖分子和氨基环醇环,由配糖键相连接。细菌对氨基糖苷类抗菌药物的耐药机制非常复杂,主要包括细胞内外电位差的改变、细菌的主动外
154 -
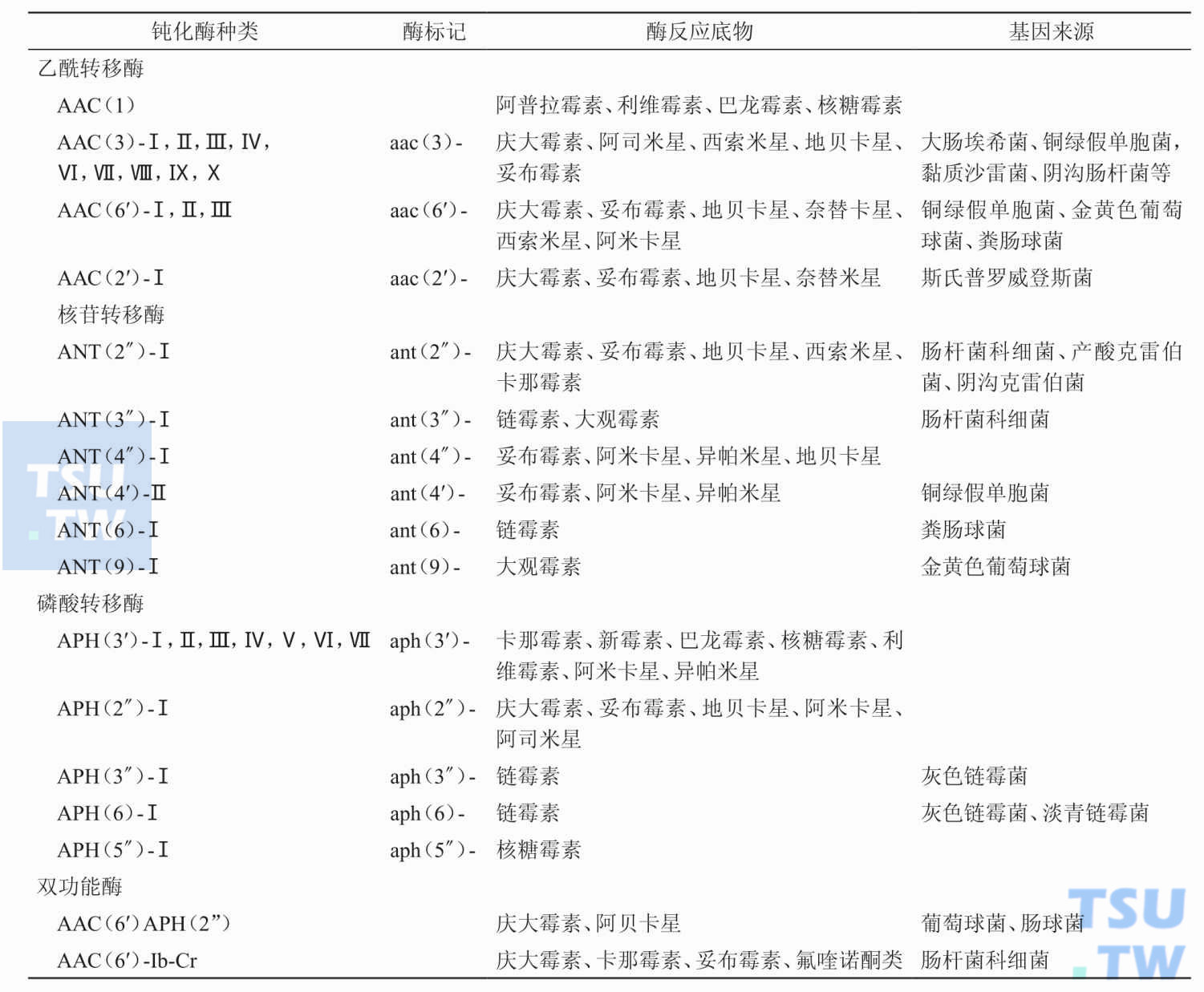
目前已知的钝化酶有哪几类?
答:目前已知有三类钝化酶,它们是:①乙酰转移酶(AAC);②磷酸转移酶(APH);③核苷转移酶(AAD或ANT)。三类酶又可按照所破坏的抗生素不同和作用点的不同而分为许多种。目前已知至少存在着30
155 -
什么是乙酰转移酶?
答:乙酰转移酶的全称应该是氨基糖苷乙酰转移酶(aminoglycoside acetyltransferases,AAC)。AAC 有4种同工酶。如AAC(1)、AAC(3)、AAC(2′)和AAC(6′)。它们主要以乙酰辅酶A作为
156 -
乙酰转移酶AAC(3)的底物谱和主要的亚型有哪些?
答:目前已经发现AAC(3)的亚型有AAC(3)-Ⅰ~AAC(3)-Ⅳ和AAC(3)-Ⅵ~Ⅹ。其中AAC(3)-Ⅰ和AAC(3)-Ⅱ临床上较为常见。AAC(3)-Ⅰ为窄谱酶,其底物包括庆大霉素、西索米星和阿司米星。临床上约30%的革
158 -
乙酰转移酶AAC(6′)的底物谱和主要的亚型有哪些?
答:AAC(6′)为广谱酶,它能修饰临床上多数氨基苷类抗生素。目前已经发现的AAC(6′)亚型有AAC(6′)-Ⅰ、AAC(6′)-Ⅱ和AAC(6′)-Ⅲ。其中AAC(6′)-Ⅰ可引起细
159 -
AAC(6′)-APH(2″)双功能钝化酶有什么特性?
答:在甲氧西林耐药的金黄色葡萄球菌、庆大霉素耐药的粪肠球菌中发现一种钝化酶蛋白,经研究发现不但具有乙酰转移酶的作用,并且还具有磷酸转移酶的作用。经编码酶的基因结构的研
160 -
乙酰转移酶AAC(2′)的底物谱和主要的亚型有哪些?
答:AAC(2′)-Ⅰa的基因分离自斯氏普罗威登斯菌(Providencia stuartii),其底物谱为庆大霉素、妥布霉素、奈替米星、地贝卡星和新霉素。有文献报道在分子杆菌属中已发现AAC(2&pr
161 -
什么是磷酸转移酶?
答:磷酸转移酶的全称应该是氨基糖苷磷酸转移酶(aminoglycoside photransferases,AHP)。目前已发现以下几种AHP磷酸转移酶。如APH(3′)、APH(2″)、APH(3″)、APH(5&Prim
162 -
磷酸转移酶APH(3′)的底物谱和主要的亚型有哪些?
答:多数APH在3′位上修饰羟基,现已经发现7种不同的APH(3′),即APH(3′)-Ⅰ~APH(3′)-Ⅶ。它们主要的底物谱是卡那霉素、新霉素、核糖霉素、巴龙霉素、利维霉素、
163 -
磷酸转移酶APH(2″)的底物谱和主要的亚型有哪些?
答:在革兰阳性菌中发现了4个APH(2″)亚型。它们是APH(2″)-Ⅰa、APH(2″)-Ⅰb、APH(2″)-Ⅰc 和APH(2″)-Ⅰd。其中编码APH(2″)-Ⅰa酶的基因位于编码AAC(6
164 -
什么是核苷转移酶?
答:核苷转移酶的全称应该是氨基糖苷核苷转移酶(aminoglycoside nucleotidyltransferases,ANT 或ADD)。目前已发现6种ANT,如ANT(2″)、ANT(3″)、ANT(4″)、ANT(4′)
165