-
核苷转移酶(ANT)的主要底物谱是什么?
答:上述6种核苷转移酶的底物谱不尽相同。其中ANT(4″)-Ⅰa和ANT(4′)-Ⅱa均可引起细菌对阿米卡星、异帕米星、妥布霉素、地贝卡星耐药,前者主要见于金黄色葡萄球菌和肠
166 -
经前述钝化酶修饰后的氨基糖苷类抗生素为何失去抗菌作用?
答:经钝化酶修饰后的氨基糖苷类可能通过下列作用而失去抗菌活性:①与未经钝化的氨基苷类竞争细菌细胞内转运系统;②与细菌的核糖体的亲和力大为降低,或不能与之相结合;③失去了干
167 -
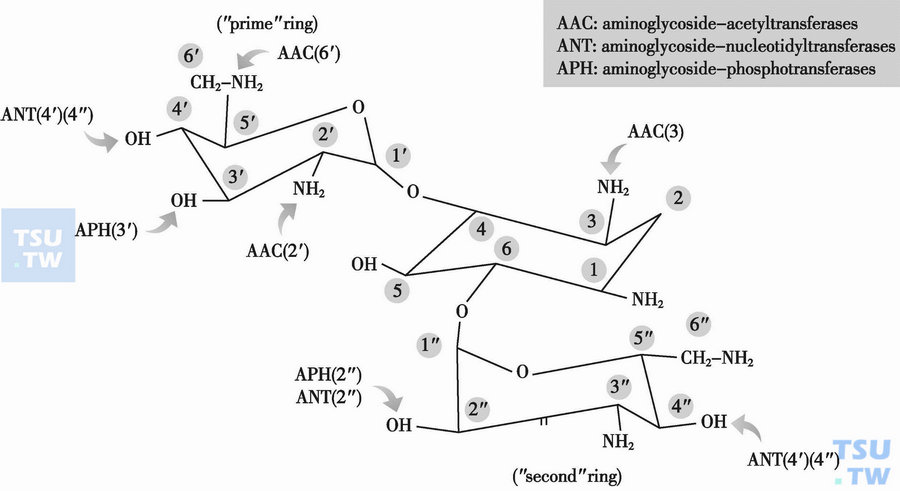
如何正确理解上述代表氨基糖苷类钝化酶的数字符号?
答:AAC即为乙酰转移酶;AAD或ANT为核苷转移酶,APH为磷酸转移酶;(1)、(3)、(6)表示主环上酶的作用位点;(2′)、(4′)、(6′)表示与主环(双脱氧链霉胺环)4位上相连接的氨基环醇环上
168 -
APH(2″)-Ⅰa和APH(2″)-Ⅰb指的是哪一类氨基苷钝化酶?
答:APH(2″)-Ⅰa和APH(2″)-Ⅰb是磷酸转移酶,意味有不同基因编码的a和b两种磷酸转移酶蛋白均能使2″位上游离的羟基发生磷酸化作用,前者使细菌对卡那霉素、庆大霉素
169 -
AAC(3)-Ⅳ和AAC(3)-Ⅶ指的是哪一类氨基糖苷钝化酶?
答:AAC(3)-Ⅳ和AAC(3)-Ⅶ就意味细菌产生的两种乙酰转移酶均可使3位上游离的氨基发生乙酰化修饰,但前者可引起细菌对妥布霉素、庆大霉素、奈替米星、地贝卡星、西索米星、阿普拉霉
170 -
编码氨基糖苷钝化酶的基因是由染色体介导还是由质粒介导的?
答:编码钝化酶的基因通常由质粒携带,其中很多还与转座子相连,并可通过接合转移或转座子转移到其他敏感菌。因此加速了这些耐药基因在菌种间的传播。如临床上约有70%的革兰阴性
171 -
不同氨基糖苷类可为同一种酶所钝化,而抗生素可为多种钝化酶所钝化?
为何不同的氨基糖苷类可为同一种酶所钝化,而同一抗生素又可为多种钝化酶所钝化?答:这是因为不同的氨基苷类抗菌药物具有相同的结合位点,如妥布霉素、庆大霉素、奈替米星、地贝卡
172 -
什么是16S rRNA甲基化酶?
答:16S rRNA甲基化酶是在2003年发现的。该酶可使细菌核糖体30S亚单位中的16S rRNA甲基化,因而使氨基苷类不能与之结合而发挥抗菌作用,导致细菌对该类抗生素耐药。此外,2006年发
173 -
除临床上已发现的氨基苷类钝化酶外,可还有其他钝化酶?
除临床上已发现的氨基苷类钝化酶外,是否还有其他种类的钝化酶?答:目前除临床上已发现的氨基苷类钝化酶外,在某些金黄色葡萄球菌、表皮葡萄球菌、D组链球菌和革兰阴性杆菌可产生
174 -
mecA基因和金黄色葡萄球菌的耐药性
自从青霉素应用于临床后的2~3年,75%的金黄色葡萄球菌即对其产生了耐药性。目前葡萄球菌中的产酶菌株约在95%左右,对青霉素均耐药。葡萄球菌对甲氧西林的耐药与细菌的β-内
175 -
近年来耐甲氧西林金黄色葡萄球菌的检出率有哪些变迁?
答:20世纪60年代欧洲首先报道耐甲氧西林金黄色葡萄球菌医院感染。到80年代耐甲氧西林的金黄色葡萄球菌和凝固酶阴性葡萄球菌已经遍布全球。目前,耐甲氧西林(苯唑西林)葡萄球菌仍
176 -
什么是mecA基因?
答:mecA基因是编码产生青霉素结合蛋白PBP2a(又称PBP2′)的结构基因,由转座子携带并整合至葡萄球菌染色体的mec部位。mec片段是葡萄球菌染色体上获得的外来片段,大约30~50kb的
177 -
PBP2a和mecA基因的关系是什么?
答:PBP2a是mecA基因编码的产物,是细菌的一种耐抗生素抑制作用的替代性靶位蛋白,分子量为76kD,它决定细菌对甲氧西林的耐药性。敏感菌株中无mecA的同源物。这就意味着在MRS菌株中
178 -
与mecA基因相关的基因有哪些?
答:mecA基因是编码青霉素结合蛋白PBP2a的结构基因,MecRI-mecl是调节基因,通过抑制mecA的转录决定PBP2a的合成水平而调节细菌的耐药程度。在细菌基因组中还存在着辅助基因femA、
179 -
什么是MRSA?
答:含有mecA基因的金黄色葡萄球菌具有对甲氧西林等β-内酰胺类抗菌药物耐药,故称甲氧西林耐药金黄色葡萄球菌,取英文名字methicillin-resistant Staphylococcus aureus的首
180