
基因表达
基因表达就是储存遗传信息的基因通过转录及翻译等步骤,产生具有一定生物功能的蛋白质的整个过程(下图)。许多蛋白质的肽链在合成后还需要经过加工修饰,如切除一定的肽段、加糖、加脂、磷酸化、羟化、与其他亚单位或因子复合等,才能成为具有完整功能的蛋白质。真核生物结构基因中与蛋白质的氨基酸顺序对应的核苷酸序列称为编码序列,其余为非编码序列。基因中与成熟mRNA顺序对应的称作外显子。基因中外显子序列是不连续的,外显子之间被内含子(又称插入序列)分隔。外显子顺序的大部分与蛋白质的氨基酸顺序是对应的,但在5'外显子和3'外显子中常存在非翻译区,分别称为5'非翻译区(5'UT)和3'非翻译区(3'UT)。
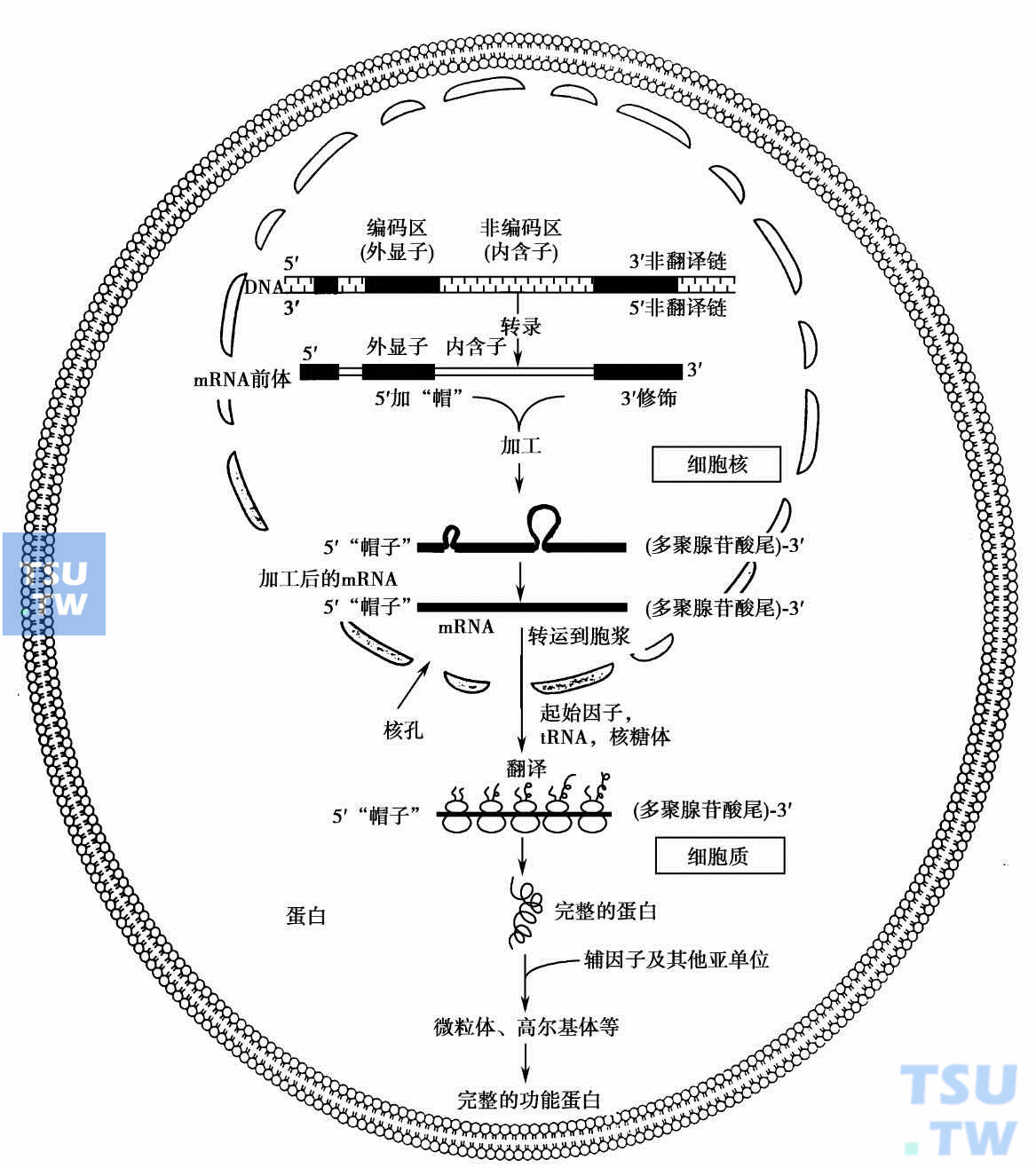
基因表达——mRNA与蛋白质的合成
转录在细胞核内进行,由RNA聚合酶以基因的一条链(模板链)为模板,按碱基配对规律,沿模板DNA链的3'→5'方向逐个接上核糖核苷酸,产生基因的初级转录物(primary transcript)。初级转录物经加工修饰形成5'“帽子”结构和3'多聚A“尾巴”,剪切掉内含子,再由外显子拼接成为成熟的mRNA(下图)。在初级转录物的剪接过程中,参加拼接的外显子可以不按其在基因组的线性分布次序拼接,内含子也可以不被切除而保留,即一个外显子或内含子是否出现在成熟的mRNA中是可以被选择的,这种剪接方式称为选择剪接。选择剪接是一种在转录后水平直接调节基因表达的调控方式,可以直接在RNA水平上调节基因的表达,也可以改变基因的表达产物,对蛋白质的功能和活性进行调节。
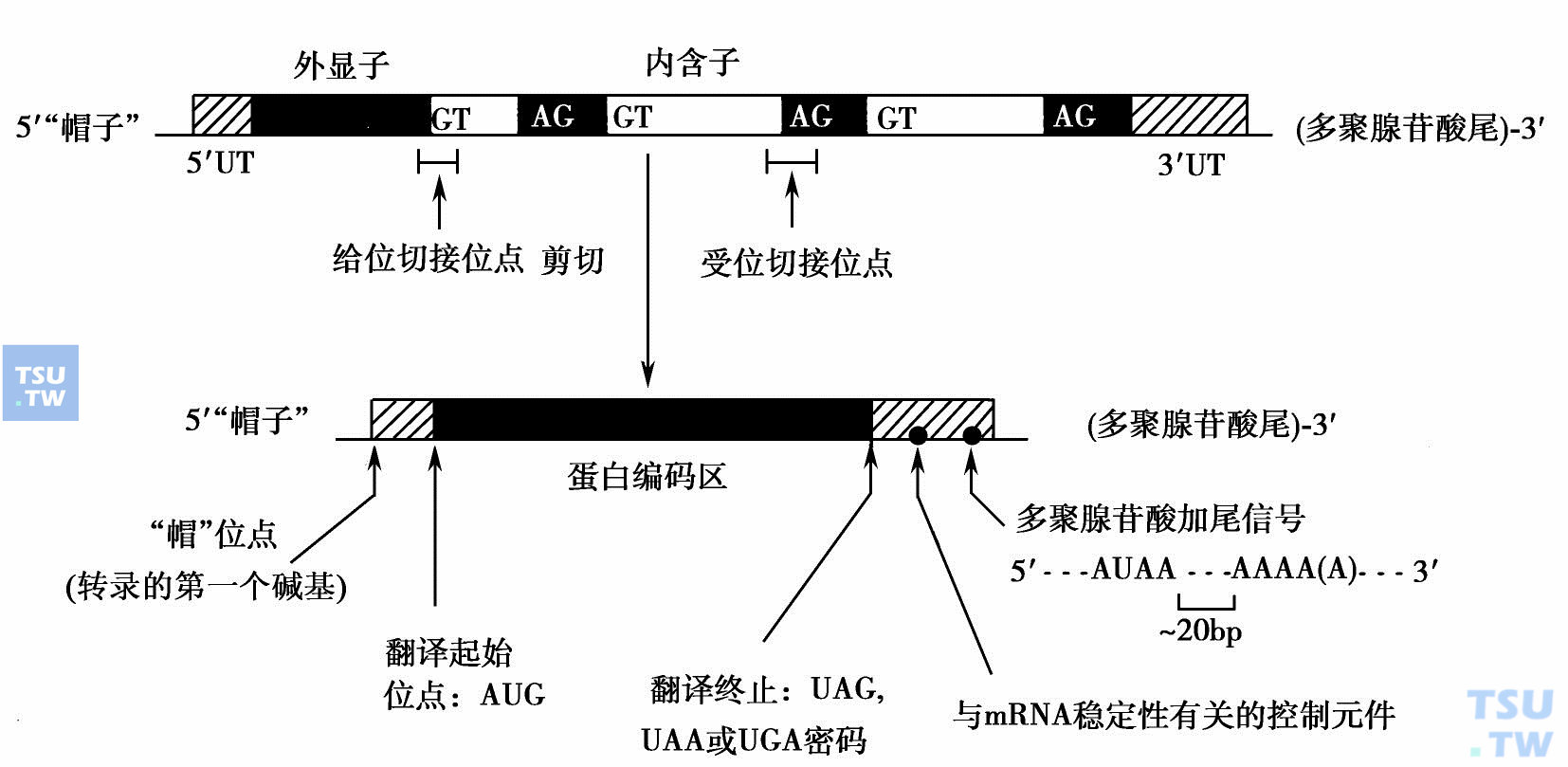
mRNA的成熟过程:5'UT:5'非翻译区;3'UT:3'非翻译区
mRNA上携带的遗传信息由三联密码子(每3个连续的核苷酸组成一个密码子)组成。4种不同的核苷酸可以组成64组密码。其中UAA、UGA和UAG为终止密码子,其他密码子均代表一定的氨基酸。AUG除代表蛋氨酸外,还是蛋白质合成的起始密码子。
成熟的mRNA被转运到细胞质中,与核糖核蛋白体结合,作为合成蛋白质的模板。由tRNA作为搬运氨基酸的特异工具,在供能物质(ATP、GTP)、无机离子(Mg+、K+)以及其他酶与蛋白因子的参与下,共同形成蛋白质生物合成体系。tRNA具有反密码子,根据碱基配对的原则识别相应的密码,但有时这种配对不典型,有所谓“不稳定碱基对”的形成。
基因表达调控
人体的任何组织或细胞都是从同一受精卵发育分化而来,具有完全相同的遗传信息,而其合成的蛋白质却不完全相同。即使在同一细胞中,不同分化发育时期或受到不同的环境或信号刺激时,所合成的蛋白质种类与数量亦常有不同。细胞中基因表达产物蛋白质浓度的调节涉及许多精细的平衡。在真核生物中,至少有7个环节可能调节细胞中某种蛋白质的终浓度;转录(即初始转录物的合成)、转录后加工、RNA从细胞核向胞质的转运、RNA的降解、多肽链合成、多肽链的修饰及加工,蛋白质降解。
转录调节是最重要的基因表达调节环节。在真核细胞内,DNA分子被严密地组织在染色质中。染色质结构对基因转录活化状态有重要影响。典型的间期核染色质可分为高度凝聚状态的异染色质(heterochromatin)和处于较为伸展状态的常染色质(euchromatin)。在常染色质中,约10%处于更为开放的伸展状态,即为具有潜在转录活性的染色质。
处于活化状态的染色质常具有的特征是:DNA低甲基化,核小体核心组蛋白高乙酰化,特异组蛋白变异体和高速泳动族蛋白(high mobility group protein,HMG)结合的增多,DNase 1高敏位点的出现(代表了局部核小体结构更为开放或核小体结构消失),转录因子结合到基因启动子区以及RNA聚合酶Ⅱ和一般转录因子的募集和前起始复合物的形成。
基因的转录调节决定性地依靠特异转录因子结合到靶基因启动子和增强子上介导。这些转录因子部分地通过募集一般转录因子和RNA聚合酶Ⅱ到靶基因行使功能。较为开放的染色质区有利于特异转录因子的结合。序列特异的DNA结合转录因子也募集辅助因子(cofactors)到基因调节区。许多辅助因子是能够通过染色质修饰促进或抑制转录的多蛋白复合物的成分。因此,特异转录活化因子的结合也促进活化染色质的形成。基因表达被染色质修饰的调节被定义为表观遗传调节(epigenetic regulation),特指不涉及DNA顺序变化的基因表达的稳定和可遗传改变。染色质修饰包括DNA甲基化以及核心组蛋白尾部的乙酰化、甲基化、磷酸化、葡糖基化、SUMO化和泛素化。这些修饰被染色质修饰酶介导。由非编码RNA介导的染色质修饰(如DNA甲基化和组蛋白甲基化)也有发现。
概括地说,基因转录调节主要靠DNA与蛋白质,以及蛋白质与蛋白质之间的相互作用来实现。这里的DNA主要指调节基因转录的顺式作用元件(cis-acting elements),蛋白质主要指反式作用因子(trans-acting factors)和辅助因子。
顺式作用元件是指与结构基因连锁并能调节基因转录的特定DNA顺序,如启动子(promoter)、增强子(enhancer)、沉默子(silencer)、基因座控制区(locus control region,LCR)和隔离子(insulator)等。
启动子是指能与转录装置(transcriptional machinery)结合并能准确有效地起始转录的特定DNA顺序。在真核生物,转录装置的主要成分除RNA聚合酶外,还必须包含一般转录因子(general transcription factors,GTFs)。RNA聚合酶Ⅱ的核心启动子(core promoters)由4个不同的元件结合组成(下图),BRE:TFⅡ(B recognition element);TATA(TATA Box);Inr(initiator element);DPE(downstream promoter element)。典型地,一个启动子仅包含这四个组件的2或3个。在基因核心启动子上游,也常包含对于启动子功能重要的DNA顺序。
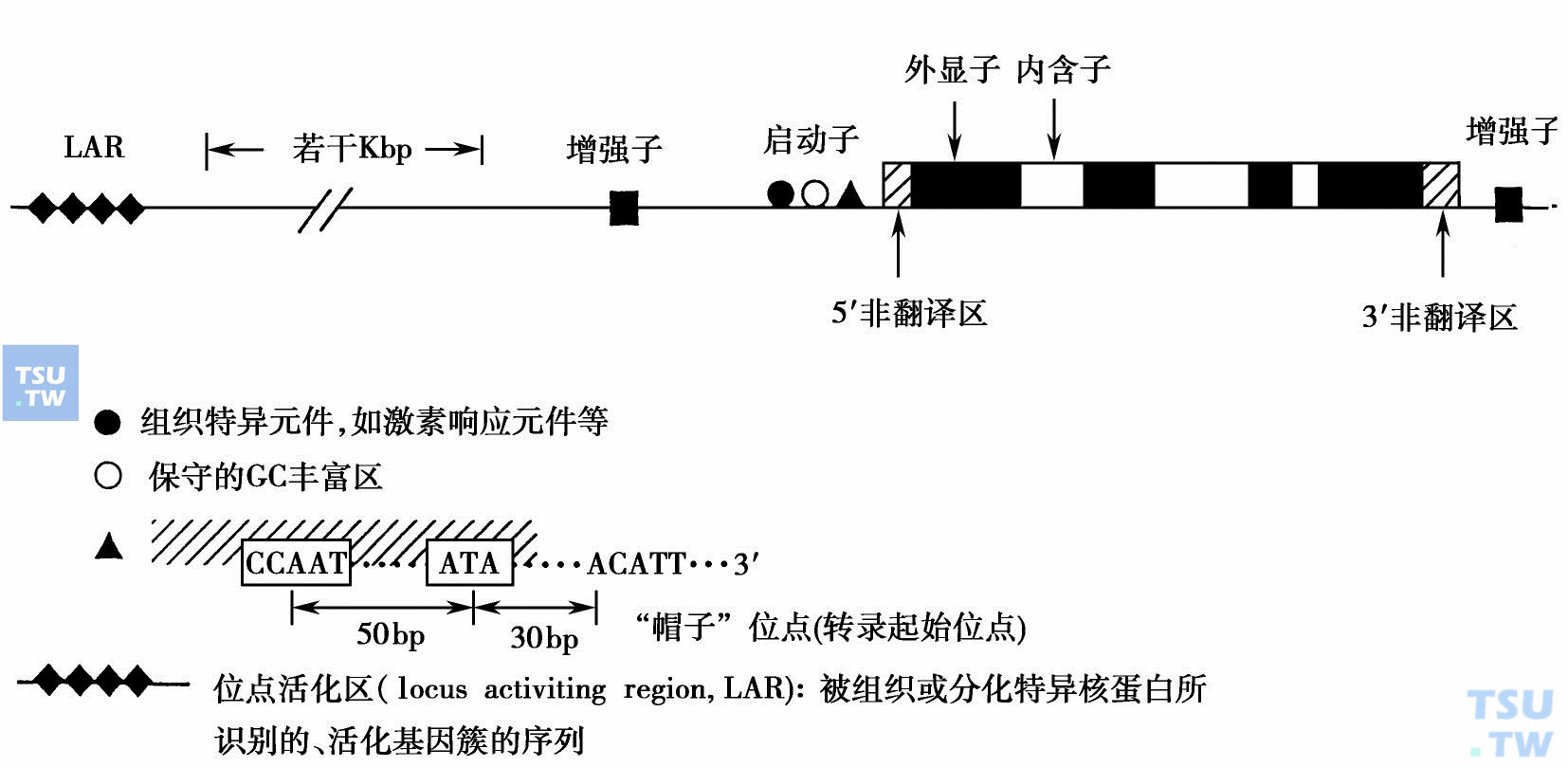
结构基因旁侧的顺式元件
在真核生物基因上游或下游常存在能增强基因转录的特异DNA顺序,称为增强子(enhancer)。它可以任一方向在基因上游或下游直至几十kb距离增强基因的转录。和启动子相似,增强子也由若干元件组成,每个元件又包含一个或几个DNA特异结合蛋白的结合位点。特异结合蛋白与增强子的特异结合可能是增强基因转录的重要环节。
基因座控制区是指能够在转基因鼠中确立连锁的基因非整合位点依赖的高水平表达能力的DNA顺序区域,它常常由若干包含DNase 1高敏位点的DNA顺序组成。特异结合蛋白与LCR的特异结合可能是增强基因转录的重要环节。
和增强子相反,沉默子是指与基因连锁的能减弱基因转录的特定DNA顺序。特异结合蛋白与沉默子的特异结合可能是减弱基因转录的重要环节。
隔离子原指当放到增强子和启动子之间时能阻断增强子作用的DNA顺序。隔离子也能抑制染色体修饰的散播。局部染色质的修饰状态可影响基因是否表达。确定的抑制性组蛋白修饰在染色质区域的散播是转录沉默(silencing)现象的核心。沉默是能够沿染色质散播的特异抑制形式,可无需每个携带特异阻遏蛋白的结合位点而关闭多重基因。隔离子能够阻断这种散播,因此隔离子能够无区别地保护基因不被活化和抑制。
一些与顺式作用元件相互作用并调节基因转录的蛋白质,称为转录调节蛋白或转录因子,又称反式作用因子。一类转录因子是大多数真核生物基因有效的和启动子特异的转录起始所必需的,称作一般转录因子,如在RNA聚合酶Ⅱ催化转录起始时所必需的若干转录因子TFⅡA、TFⅡB、TFⅡD、TFⅡE、 TFⅡF和FⅡH等。另一类为对特异基因转录起调节作用的转录因子。一些蛋白因子并不直接与DNA结合,而是通过与转录因子的相互作用起到调节基因转录的功能,称为辅助因子。
转录因子至少有两个功能:特异地结合到DNA上及调节转录启动。转录因子的一个重要特征是它们常具有标准的组成和结构,即motif(模体,也称作基序)。这些模体对于转录因子特异识别和结合到DNA上具有关键性作用。真核生物中的大部分转录因子包含4类模体中的一类:同源结构域(homeodomains),包含了螺旋-转折-螺旋(helix-turn-helix,HTH)结构;锌指(Zn-finger或Zn-binding domains),是某些蛋白质分子中由若干保守的氨基酸残基同锌离子结合形成的相对独立的“指”状四面体结构;亮氨酸拉链(leucine zipper);螺旋-环-螺旋(helix-loophelix,HLH)。少数转录因子不属于上述四种类型的任何一类。
转录后调控对于基因表达也是重要的。特别值得提及的是近几年发现的一类具有重要调控功能的微小RNA(microRNA)。microRNA是近几年发现的非编码RNA(non-coding RNAs)的主要种类,长度在19~25个核苷酸。到2009年3月在Sanger miRBase Sequence database ver.12. 0上共收录了人类microRNA 706个。miRNA已被公认为是多细胞生物基因表达转录后水平调控(通过结合到与靶mRNA互补的顺序,使靶mRNA剪切或翻译抑制)的一种普遍方式,并可能调节30%以上的蛋白质基因表达。microRNA也已被证明参与许多重要的生命过程,并涉及多种疾病,特别是癌症。
基因突变
基因在内外环境的许多因素影响下都可能发展成突变,其中有些突变推动着生物的进化,有些为中性突变,也有一些突变会导致基因产物蛋白质结构的变化或不能正常表达而造成不良后果。涉及基因结构的突变可分类为点突变、插入、缺失、重排和动态突变。
点突变指基因中的某一个碱基为另一种碱基取代。如取代后不改变密码子的意义(由于密码的简并性),称为同义突变(samesense mutation)。如果这种取代导致蛋白质分子中相应位置氨基酸改变则称为错义突变(missense mutation)。错义突变后果的严重性取决于突变的位置及其对蛋白质功能影响的程度。无意义突变(nonsense mutation)是指在基因编码区发生点突变后形成了不应有的终止密码,导致肽链合成的提前终止。有时点突变如果发生在mRNA剪接位点上,也会造成内含子不能被正确剪接,影响mRNA的转运、稳定性,或合成出异常肽链。如果突变发生在基因启动子或增强子重要部位,也会影响基因转录。
DNA大片段的缺失或插入可以造成整个基因的丢失、基因结构的改变、基因功能的丧失。发生于基因编码序列中非3整数倍碱基的插入或缺失,可引起阅读框架的位移。从移码处开始产生氨基酸序列完全不同的肽链,造成提前终止或形成无功能或不稳定的蛋白质。例如,很大一部分α地中海贫血患者是由于α珠蛋白基因缺失所致。凝血因子Ⅷ基因14外显子中2~4kb DNA片段的插入则导致甲型血友病。
染色体易位可造成两个原来不在同一条染色体上的基因间发生重排。如90%~95%的慢性粒细胞白血病(CML)患者具有Ph染色体,t(9;22)(q34. 1;q11. 21)易位。染色体易位使原来分别位于9号染色体上的abl基因与22号染色体上的bcr区域重排,产生abl/bcr融合基因。这一融合基因可转录特异的mRNA,并合成肿瘤特异的蛋白质。此外,急性淋巴细胞白血病、B细胞慢性淋巴细胞白血病、急性T细胞白血病等血液系统肿瘤均与基因重排有关。
在一些基因编码区或其旁侧存在着串联重复排列三核苷酸序列。这些序列的突变方式是三核苷酸序列重复次数的改变。其拷贝数的增加有一渐进和积累的过程,突变频率与拷贝数有关,突变体与其上一代的突变频率不同,故称动态突变。这类突变导致的疾病有不同程度的中间型表型,表现为遗传学上的不完全显性和“早现”(发病年龄逐代提前,临床表现逐代加重)现象。这类突变可改变基因的转录活性与mRMA稳定性,或表达异常肽链。强直性肌营养不良、亨廷顿舞蹈病等神经肌肉性疾病均为动态突变所致。
