
皮肤组织是大量的化学信号物质的作用靶位,例如,毛囊和皮脂腺是雄性激素作用的靶位,黑素细胞受垂体的多肽激素的影响。传统认为,激素物质是特殊的无管腺体分泌到血流中的一种物质。目前,皮肤和其他组织已不再被简单地认为只是这些物质作用的靶位,皮肤等组织不但能接受,同时还参与形成和释放信号物质,从而成为这些信号网络中的—个重要成员。因此从广义的角度来看,皮肤组织不但是激素作用的靶位,同时也是激素的生产者,具有内分泌的意义。
受体是细胞表面或亚细胞组分中的一种分子,可以识别并特异地与有生物活性的化学信号物质(配体)结合,从而激活或启动一系列生物化学反应,最后导致该信号物质特定的生物效应。
根据受体在细胞中的位置,可以分为细胞表面受体(水溶性多肽激素)和胞内受体(如甾类激素等)。它们参与不同的信号传导途径。
甾类激素的受体存在于细胞质中。甾类激素受体的氨基酸数目从400到900不等,都具有一个高度保守的富含半胱氨酸的区域,它由70-80个氨基酸组成二个“锌指结构”的重复单位。不同甾类受体的共同特点是有三个功能区:富含Cys、具有锌指结构的DNA结合区(C区);C端的激素结合区(E区)及N端的受体调节区(A/B区)。A/B区具有一个转录激活结构(另—个在E区),而且决定启动子专一性和细胞专一性,即它在选择激活不同靶基因,决定激素多样性上有重要意义。E区除激素结合域外还有与转录激活、受体二聚化及抑制蛋白结合的结构域。
糖皮质激素受体
糖皮质激素受体( glucocorticoid receptor.GR),特异的糖皮质激素受体在人类的表皮、真皮、培养的角质形成细胞和成纤维细胞中已得到证实。它广泛地分布于体内各部位:肝、脾、肾小管、胎盘、外周血细胞及皮肤,受体主要位于细胞质中。与受体的结合能力表皮要高于真皮。糖皮质激素对靶细胞的治疗效应必须通过特异性受体的介导来实现。GR和甲状腺激素受体、维A酸受体、性激素受体等均属于核激素受体超家族的成员,这类受体是一种配体依赖的转录活化因子。GR包括GRa和GRp,两者为GR基因同一转录产物通过不同的剪切方式剪切的结果。
GR是由多肽链组成的酸性蛋白质分子,已查明其完整的氨基酸排列。其等电点为5.8,分子量为40-95kD,动力黏度为3.5-6.0 nmo具有三个独立的功能区:激素结合部(在氨基端)、DNA结合区(在分子的中端)和抗原决定簇(位于羧基端)。糖皮质激素受体的基因定位于第5号染色体上,通过磷酸化作用而转化为有活性形式,脱磷酸则失去活性。
性激素受体
性激素的生理作用主要为促进性器官的生长发育,促进蛋白质的合成等;性激素的这些作用均是通过性激素受体来介导的。性激素(雄激素、雌激素及孕激素)受体均为核受体超家族成员。皮肤及其附属器是性激素的靶器官之一。
性激素受体是一类可溶的酸性糖蛋白分子,很易变性,如受热、蛋白酶和不适当的pH均可减低其活性。在靶细胞内,性激素具有活化与非活化两种状态,并且有单体、二聚体和四聚体三种形式。它们有三型结合点:Ⅰ型为经典型受体,Ⅱ型和Ⅲ型作用不明。靶组织中性激素受体单体分子量:雌性激素受体70 kD,雄性激素受体66 kD,孕激素A亚基70-94 kD、B亚基100-120 kD。不同方法所测得的分子量可能不完全相同,这可能与受体所处状态、型别及受体的加工与修饰有关。性激素受体有三个功能区:羧基末端含有性激素结合区和—个二聚化区,中央区形成两个锌指结构为DNA结合区,氨基末端为受体的高变区决定其种类特异性。
雄性激素受体
皮脂腺、一些毛囊和顶泌汗腺是雄性激素的特异靶位。一些皮肤疾病,如寻常痤疮、化脓性汗腺炎、男性脱发和妇女男性型脱发等疾病是雄性激素依赖性的。
雄性激素受体是雄性激素依赖的毛发生长所必需的,因为口服抗雄性激素药物能抑制体毛,而且在一些遗传性疾病中,如XY型合子的睾丸女性化患者,发展为女性表现型,因为他们缺乏雄性激素受体,即使在血浆中有高出正常2-4倍的睾酮,仍然缺乏阴毛。在这种个体中,一些雄性激素依赖的地方,如阴阜部位,皮肤似乎缺乏5α-还原酶。
雌性激素受体
和孕激素受体有关雌性激素对皮肤的作用尚不明确。迄今能证实雌性激素在维持真皮和骨胶原以及增加基础物质的合成方面具有作用。给予药理剂量的雌性激素能抑制皮脂腺的分泌。已发现无论女性或男性痤疮患者,其雌性激素受体都较正常水平高。已发现在皮肤细胞胞质中有孕激素受体存在,胸部受体水平最高,阴阜部最低。但是,孕激素对皮肤的作用不清楚。
维A酸受体
(retinoic acidreceptors.RARs),维A酸( retinoicacid.RA)类化合物是一组与维生素A结构上类似的具有生物学活性的化合物,在脊椎动物的发育、细胞分化和维持体内平衡中发挥广泛的效应。人工合成维A酸及其内源性类似物全反式RA在活体内外对表皮角质形成细胞的功能均有极大的影响,RA与其核受体结合发挥生物学作用,在皮肤科RA具有广泛而重要的作用,在治疗诸如银屑病、皮肤肿瘤、角化性皮肤疾病、色素沉着性疾病以及寻常痤疮等疾病中,RA具有明确而重要的疗效。
RA作用的介导与两类特异性结合RA的细胞内蛋白相关:一类是RA受体RAR和RXR,属于核受体超家族。RAR与RXR之间组成的异二聚体或同二聚体可结合于不同靶基因的启动子区域。另一类是胞质RA结含蛋白(cytoplasonia retinoic acid bindulg proteui,CRABP),CRABP是小分子量蛋白,发现于所有的脊椎动物并且在物种间高度保守。
维A酸结合蛋白
在脊椎动物,CRABP包括两类同源蛋白CRABP Ⅰ和CRABP Ⅱ。在成人,CRABP Ⅰ几乎存在于所有细胞中,而CRABP Ⅱ则主要在皮肤、子宫、卵巢和脉络膜丛中表达。CRABP在胞质中保护和转运其配体并使之可溶。
核受体
维A酸受体与维A酸X受体在靶细胞核内,维A酸分子与其受体结合发挥作用。维A酸受体有两类,即RARs和维A酸X受体(reUnoid X receptors,RXR)。其中,RXR不仅与RAR形成异二聚体,还与维生素D受体、甲状腺素T3受体、孤儿受体等其他核受体形成异二聚体,激活相应的信号传导。RAR与其他核受体竞争同RXR形成异二聚体,竞争力取决于各自的表达水平及与RXR的亲和力。
维A酸受体分子结构基本相同,为一条肽链,从N末端到C末端分为A-F6个区。C区为DNA结合区,对与目标基因上游起动区DNA序列的识别和结合至关重要:E区为配体结合区,具有与配体特异性结合并传导配体诱导的交叉激活功能,且提供受体间的相互作用面,与受体二聚体的形成有关。维A酸与RAR的配体结合区相结合,通过RXR与RAR异二聚体形式,以RAR的DNA结合区与基因启动区的维A酸反应元件结合,激活或抑制靶基因转录,从而调节基因表达。
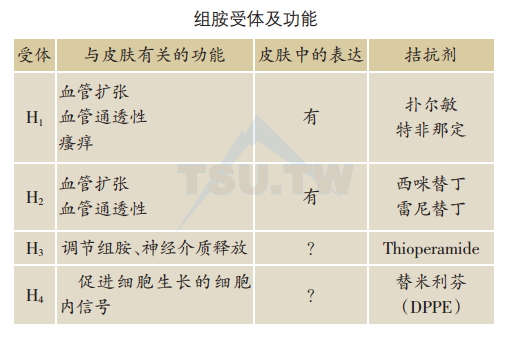
组胺主要通过组胺受体发挥其生理功能,如组胺H2受体与血管的通透性、血管扩张和平滑肌的收缩有关;组胺H2受体参与抑制嗜碱性粒细胞、中性粒细胞和淋巴细胞介质的释放,但能刺激胃壁细胞的分泌;通过组胺H3受体调节自身及其他神经递质的释放。最近为了寻找新的G蛋白偶联受体,在基因库内筛选到了一个与组胺H3受体cDNA同源性很高的基因序列,经克隆表达后发现该受体能与组胺结合,且和组胺受体的配体结合时显示独特的药理学性质,确认为不同于组胺H1、H2、H3受体的新的组胺受体,命名为组胺H4受体,其生理作用尚未明确。组胺受体及功能见下表。
其他
血管内皮细胞生长因子受体
vascular endothelial grovrth factor receptors,VECFR:血管内皮细胞生长因子(VECF)通过血管内皮细胞特异性受体VECFR介导血管内皮细胞的增殖、微血管增生及增强血管通透性。目前已经发现的VECFR可分为两大类:酪氨酸激酶受体和非酪氨酸激酶受体,前者包括VECFR-1、VECFR-2和VECFR-3,后者包括神经纤毛蛋白-1 (neumpilin 1,NRP-1)和神经纤毛蛋白-2(neuropilin 2.NRP-2)。VEGFR-1是最早发现的VECFR.是相对分子质量为180 kD的糖蛋白,与VECF、胎盘生长因子(placema groWth factor,PICF)和VECF-2有较高的亲和力。在生理情况下,主要起负向调控VEGF生物学作用,而在某些病理情况下发挥正向调控VEGF的作用。
新近研究表明,在正常人表皮中VEGFR-1、VEG-FR-2、VECFR-3及NRP-1和NRP-2均有不同程度的低表达。VECF可通过VECFR-2促进角质形成细胞的增殖、迁移和黏附。VEGFR及NRP可能在正常表皮角质形成细胞的生物学行为中发挥重要作用。郑敏等发现银屑病患者皮损部位的表皮中表达VEGFR-1、VECFR-2、VECFR-3等均有明显上调而且与疾病进展有关。近年较多的研究证据表明黑素瘤细胞本身高表达VECFR,外源性的VECF通过作用于黑素瘤细胞的VECFR -2激活MEK/MAPK及PI3/Akt磷酸化途径,促进肿瘤细胞增殖及向细胞外基质浸润,而VECFR酪氨酸激酶抑制剂能够抑制肿瘤细胞的增殖及浸润。皮肤外伤后局部炎症能够促进角质形成细胞分泌VECF,从而能通过旁分泌作用于血管内皮细胞上的VECFR,促进血管增生及通透性增强,促进成纤维细胞和角质形成细胞增殖,还可能通过自分泌的作用促进上皮细胞增殖及伤口愈合。
Toll样受体
ToIl-like receptor,TLR:TLR是新近发现的一类天然免疫受体,许多研究表明TLR的适度激活能够启动天然免疫清除病原茵,但同时发现TLR的过度表达也可加重疾病的发生发展。TLR在生物界广泛存在,它们在天然免疫中发挥重要的抗感染免疫功能。
TLR是表达于免疫细胞表面的一类Ⅰ型跨膜糖蛋白,可分为胞外区、跨膜区、胞质区(又称为Toll/L-1R同源区)。其胞外区富含亮氨酸重复序列,参与对病原微生物或其产物的识别。不同TLR胞外区氨基酸组成的差异性决定了不同的TLR识别不同的配体。如TLR4识别脂多糖与热休克蛋白,TLR3识别病毒的双链RNA,TLR2识别革兰阳性茵的细胞壁成分——肽聚糖,TLR5识别鞭毛蛋白,TLR9识别细菌中未甲基化的CpCDNA成分,TLR6/TIR2异物二聚体识别二酰化脂蛋白,TLRI/TLR2异物二聚体识别三酰化脂蛋白,TLR8识别单链RNA,TLR7识别人工合成的咪唑并喹啉类化合物,如咪喹莫特和瑞喹莫特。新近于人体肾脏组织中发现的TLRII主要识别大肠埃希菌类的非致病性细菌,其识别的具体成分还有待研究。
研究表明正常的角质形成细胞表达TLR1、2和5,其中TLR1、2表达于表皮各层,并且TLR2更侧重表达于表皮的基底层,而TLR5只表达于表皮的基底层,这表明TIR的表达可能随细胞成熟程度不同而改变。
TLR参与皮肤抗细菌免疫,在皮肤组织中,TLR经多种途径参与抗细菌免疫,其中最突出的生物学功能即是促进细胞因子的合成与释放,引发炎症反应。皮肤中的单核巨噬细胞、树突状细胞等通过膜表面表达的TLR感受入侵病原体的病原体相关分子类型(pathogen-associated molecular pattem,PAMP)刺激后,经由胞内信号传导通路,最终合成并释放IL-1、IL-6、IL-8、IL-2、TNF-α及IFN-γ等细胞因子,引起粒细胞、巨噬细胞趋化聚集,毛细血管通透性增高,淋巴细胞浸润等炎症反应,从而发挥早期免疫应答的效应。病毒感染可通过TLR7、TLR9诱导Ⅰ型IFN(IFNα/β),发挥抗病毒作用。某些局部免疫调节剂即利用TIR信号途径发挥抗病毒免疫。例如咪喹莫特在临床上用来治疗包括尖锐湿疣在内的多种病毒性皮肤病,并取得了较好的疗效。有研究表明咪喹莫特主要通过TLR7介导Ⅰ型IFN的生成,从而发挥抗病毒效应。
有研究表明,TLRs参与特应性皮炎、痤疮、麻风、艾滋病、念珠菌病、单纯疱疹等发病过程。对TIR的深入研究,有利于揭示许多包括皮肤病在内的疾病的发病机制,并为开发治疗这些疾病的有效药物提供新思路。
