-
什么是非特异性整合?
答:非特异性整合指的是耐药基因盒并不结合于attC和attI交换位点,而结合于整合子的非特异性交换位点,亦称第二位点。这种整合形式发生的频率很低,但只要发生这种整合,整合上的基因
92 -
基因盒的表达与什么有关?
答:耐药基因盒的表达依赖位于整合子5′保守端的启动子,耐药基因才得以表达。基因盒的表达不仅依赖启动子的强弱,而且与启动子的距离远近有关。对于特异性整合,越靠近启动子
93 -
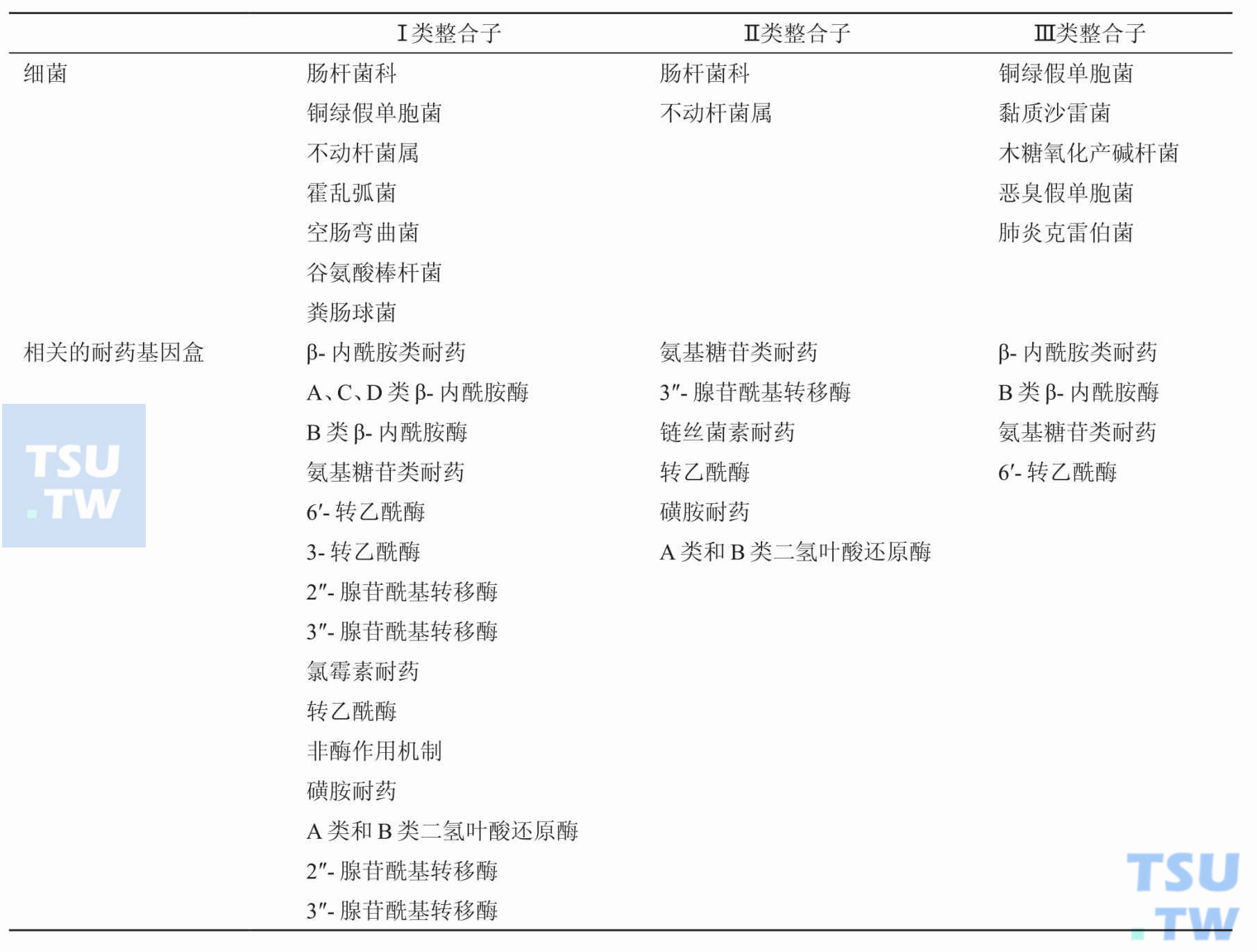
目前已知和细菌耐药性有关的整合子有哪几类?
答:根据整合酶基因序列不同,目前发现的整合子分为5类,其中Ⅰ类、Ⅱ类和Ⅲ类整合子已被证明与细菌耐药性有关。与细菌耐药有关的从临床株中发现的整合子大多属于Ⅰ类整合子。Ⅰ
95 -
Ⅰ类整合子有哪些基本结构?哪些临床分离菌中最常见Ⅰ类整合子?
答:从临床株中发现的整合子以Ⅰ类整合子最为常见,它与Tn21转座子家族相关,其整合酶是含有337个氨基酸的蛋白。Ⅰ类整合子具有整合子的基本结构:5′保守末端有编码整合酶的
97 -
Ⅱ类整合子有哪些基本结构?哪些临床分离菌中最常见Ⅱ类整合子?
答:Ⅱ类整合子则与Tn7转座子家族相关,intI 2基因产物与intI 1的产物有46%的同源性,其3′保守端有5个tns基因和Tn7的转移有关。
98 -
Ⅲ类整合子有哪些基本结构?哪些临床分离菌中最常见Ⅲ类整合子?
答:Ⅲ类整合子是在耐碳青霉烯类抗生素的黏质沙雷菌的质粒上发现的,另外还在肺炎克雷伯菌、假单胞菌、产碱杆菌等革兰阴性菌中分离出Ⅲ类整合子,目前只发现碳青霉烯类耐药基因位
99 -
什么是超级整合子?
(14) 什么是超级整合子?答:指在霍乱弧菌染色体上存在与基因盒attC位点相似的序列。这些序列被称为霍乱弧菌重复序列(Vibrio cholerae repeat,VCR)。VCR基因序列中某些基因编码毒素
100 -
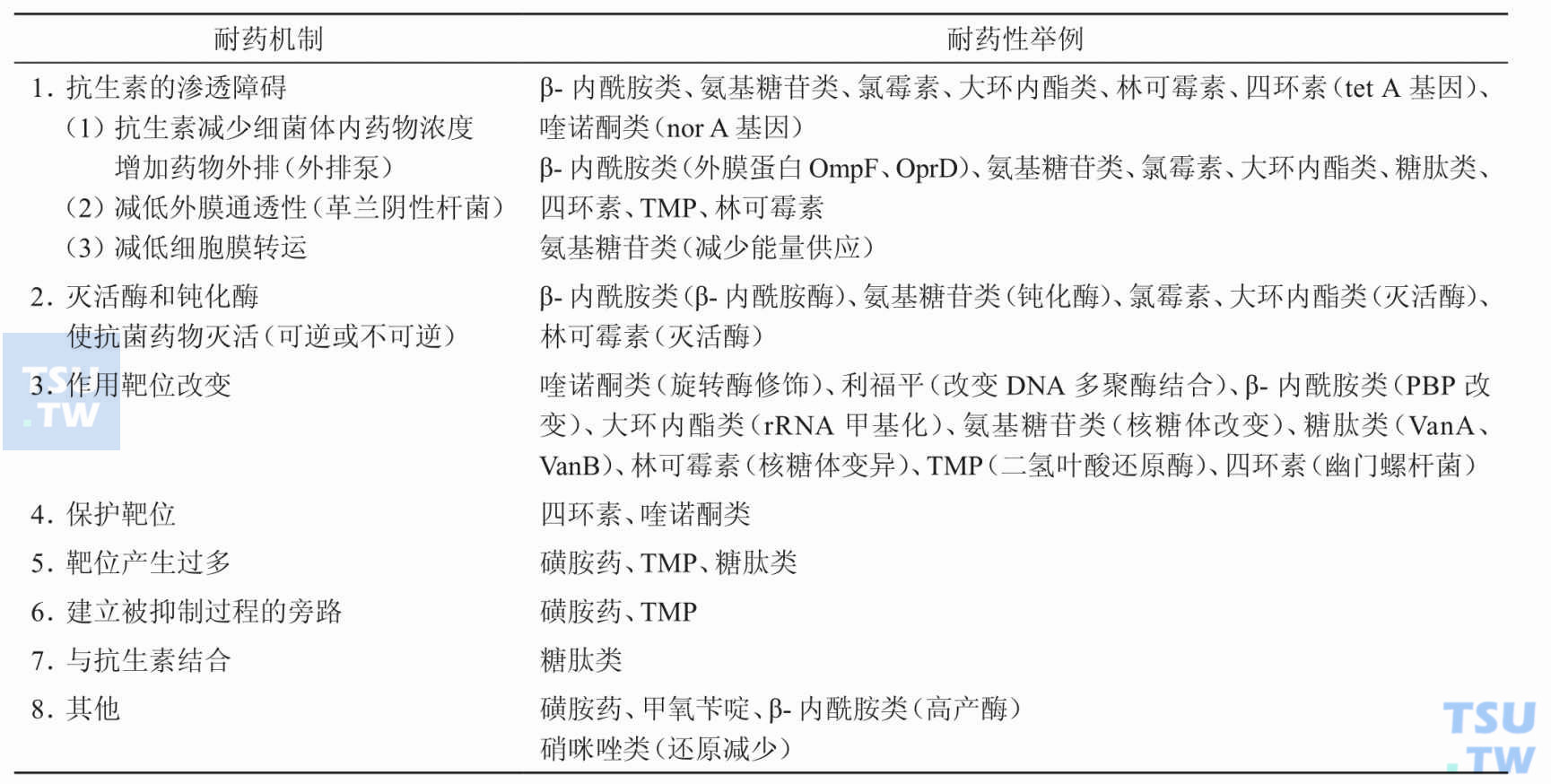
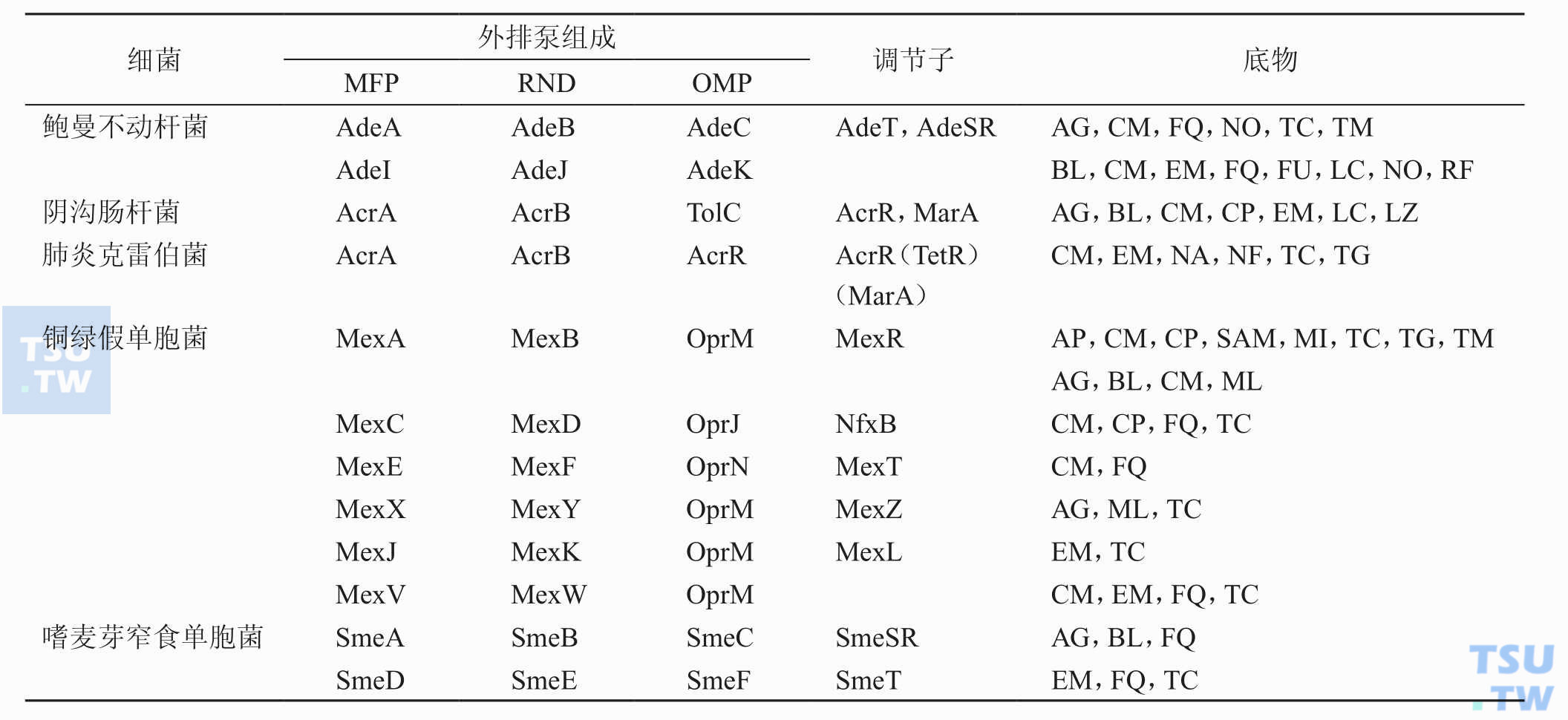
引起细菌耐药性的机制有哪些?
答:细菌大凡可通过以下几个途径来达到对抗菌药物的耐药性(下表):①灭活酶或钝化酶的产生;②抗生素的渗透障碍;③药物作用靶位改变;④产生靶位保护蛋白;⑤细菌还可增加对抗菌药物拮抗
102 -
为什么灭活酶或钝化酶的产生可引起细菌的耐药性?
答:细菌可通过耐药基因编码产生破坏抗生素的活性基团使之失去抗菌作用的酶,使药物在作用于菌体前即被破坏或失效。如细菌可产生β-内酰胺酶,使β-内酰胺类抗生素的活性
103 -
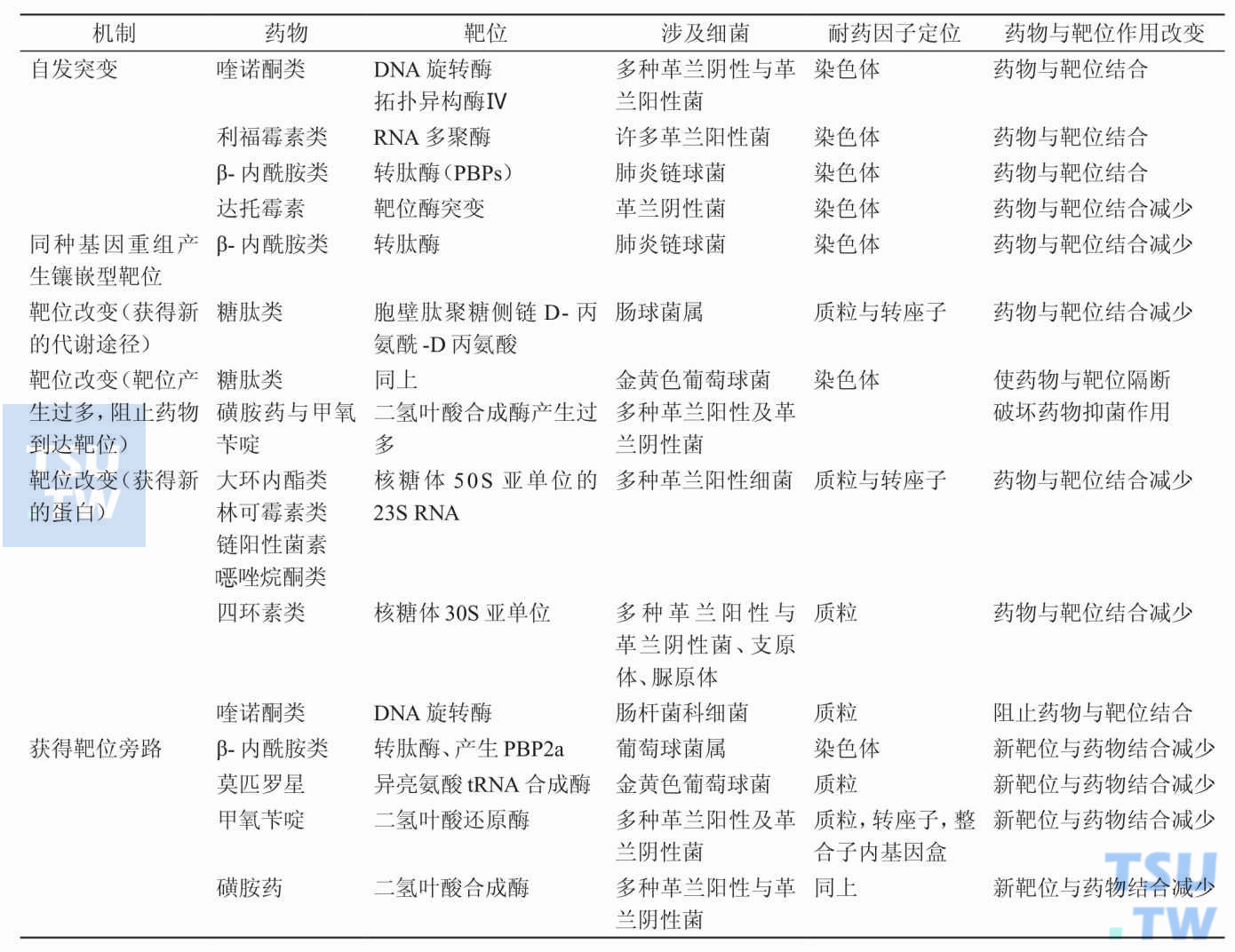
为什么靶位的改变可引起细菌的耐药性?
答:细菌可改变抗生素作用靶位的结构,使其不能与作用靶位结合发挥其抗菌活性。如细菌可改变抗生素与核糖体的结合部位而导致四环素、大环内酯类、林可霉素类与氨基苷类等抗菌药
105