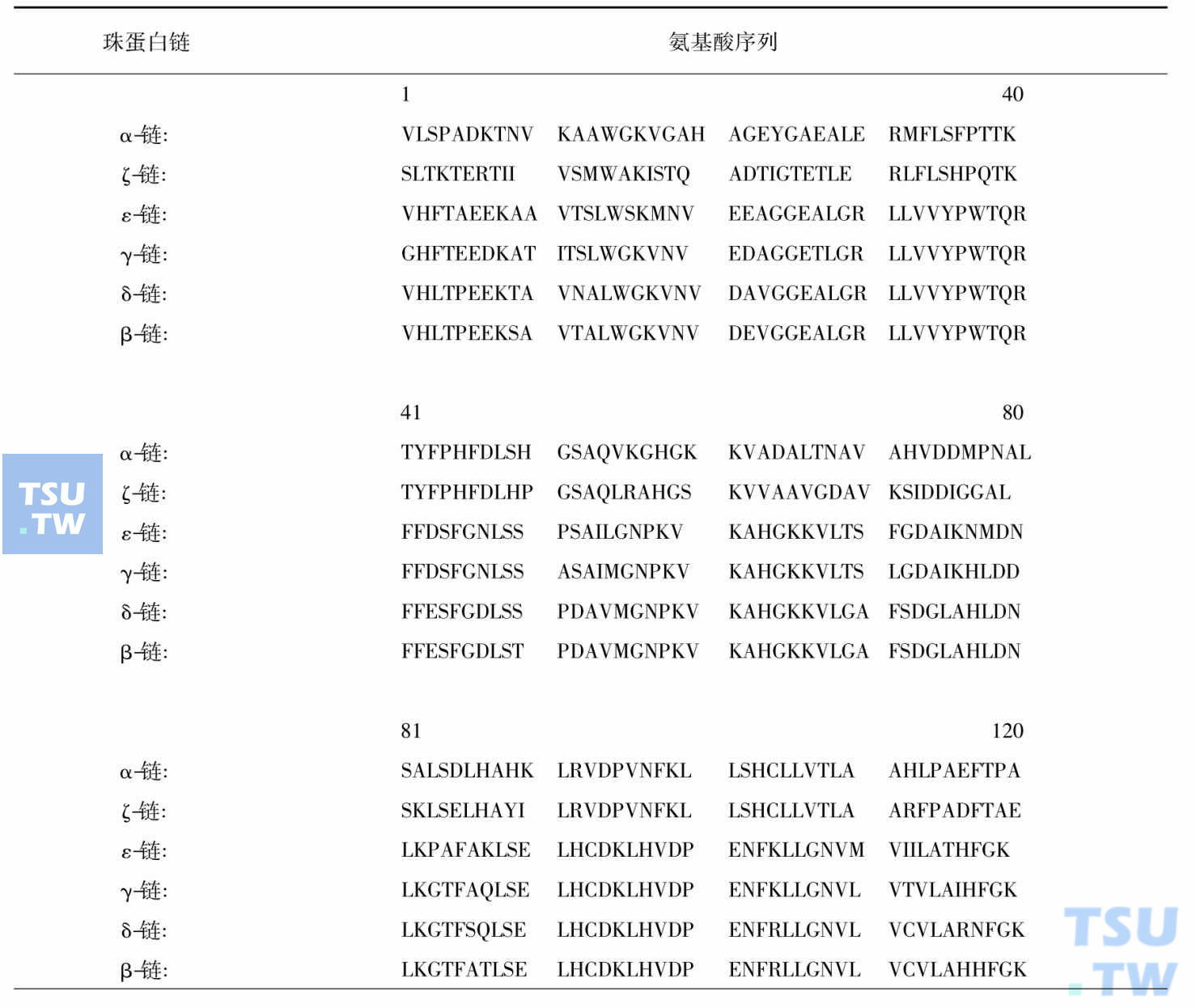
由上几篇所述不同发育阶段和不同场所合成的α类和β类珠蛋白肽链组成血红蛋白四聚体至少可分为6种(下表),胚胎型三种:HbGowerⅠ(ζ2ε2)、HbGowerⅡ(α2ε2)、HbPortland(ζ2γ2);胎儿型一种:HbF(α2γ2);成人型两种:HbA(α2β2)、HbA2(α2δ2)。每种血红蛋白都含有两条α类珠蛋白肽链和两条β类珠蛋白肽链,呈现α类珠蛋白基因与β类珠蛋白基因的表达始终保持平衡,如果这一平衡失调就导致α地中海贫血和β地中海贫血症。
人发育期血红蛋白的组成
血红蛋白A(HbA)是成人中主要的血红蛋白,由一对α链和一对β链组成(α2β2),其分子直径为50Å,体积约为64×55×50Å,分子量为65kD,一个血红蛋白分子含4个原子铁(0. 346%W/W)。HbA在氧合和脱氧时的吸收光谱不同(下图),氧合Hb在540、580nm处各有一个吸收峰,而脱氧Hb只在560nm处有一个吸收峰。在新生儿中HbA占所有血红蛋白的20%左右,出生后快速增加,出生6个月后成为血红蛋白中的主要成分,占所有血红蛋白的90%以上。这种血红蛋白是目前所知最多的一个,关于它的结构与功能关系的研究极大地丰富了对蛋白质的认识。本章主要是以它为中心进行讨论。
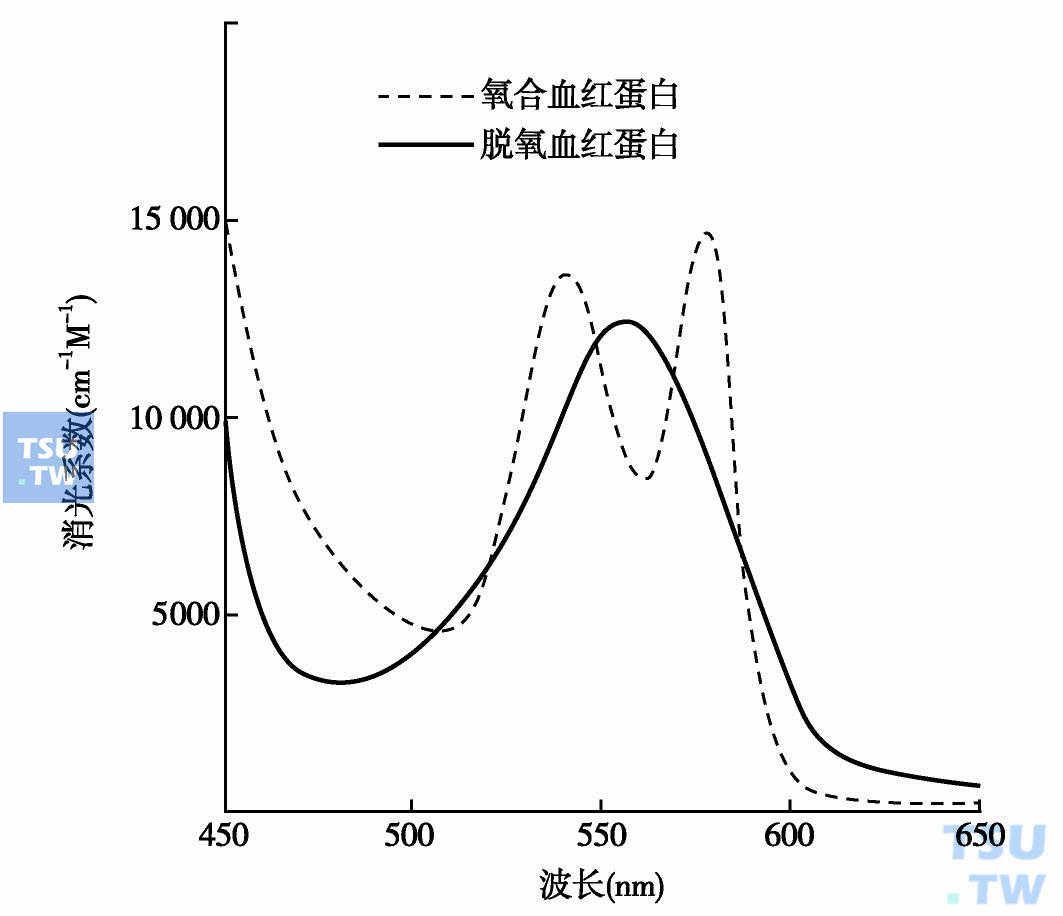
氧合和脱氧血红蛋白的可见吸收光谱
血红蛋白A2(HbA2)是正常人红细胞中的次要血红蛋白成分,它是由一对α珠蛋白链和一对δ珠蛋白链组成(α2δ2)。在整个正常人血红蛋白中,HbA2的含量少,出生6个月后,HbA2仅占所有血红蛋白总量的2%~2. 5%,但它在各红细胞中是均匀分布的。HbA(α2β2)和HbA2的差别在于β类亚基,在HbA为β亚基,而在HbA2为δ亚基。在化学结构方面,δ珠蛋白链和β珠蛋白链具有高度同源性,均含有146个氨基酸残基,其顺序见下表。二者的N-端氨基酸均为缬氨酸(Val),C-末端氨基酸均为组氨酸(His),在146个氨基酸残基中只有表中的10个不同。
这10个不同残基中的8个是中性氨基酸的互换,然而β22Glu→δ22Ala及β116His→δ116Arg的改变,使得δ珠蛋白链的正电荷多于β珠蛋白链,从而造成HbA2的等电点升高。
δ珠蛋白和β珠蛋白链氨基酸残基顺序
在物理化性质方面,HbA2与HbA有些不同,具有自己的特点。HbA2的等电点较高,所以碱性电泳时向HbA的阴极移动。它的层析行为也不同于HbA,DEAE纤维素柱层析中,HbA2先于HbA被洗脱下来。在抗原性方面,HbA2比HbA强。Nanney等发现,HbA2和它的氧化变性产物——高铁血色原A2(hemichrome A2),与红细胞膜,特别是与红细胞膜蛋白区带3的结合力大于HbA和高铁血色原A与红细胞膜结合力。氧合HbA2在磷酸缓冲液中的溶解度稍低于氧合HbA。尽管HbA和HbA2性质上有这些不同,但是二者在生理功能上的特性几乎是一样的,例如HbA2的氧亲和力、与2,3-二磷酸甘油酸(2,3-DPG)结合能力、Bohr效应和协同效应与HbA没有显著差别。在成人体内由于HbA2含量低,行使生理功能的主要是HbA。虽然HbA2在生理上并不重要,但却没有在漫长的进化过程中被淘汰,说明它可能还具有未知的重要功能,如1993年Poillon发现HbA2具有抑制红细胞内脱氧HbS聚合作用,通过增加HbA2合成量可以保护大部分含有HbS的红细胞免于镰刀形化损伤,这有希望为治疗镰刀型贫血病提供一条新途径。此外,还有人证明HbA2能增加HbA的热稳定性。
血红蛋白F(HbF)又称胎儿血红蛋白,由一对α链和一对γ链组成(α2γ2),是胎儿第2个月后和新生儿血液中的主要血红蛋白。脐带血中HbF的含量占总血红蛋白的70%~80%,出生后很快减少,至6个月后,绝大部分被HbA替代。成人中HbF约占总Hb的1%左右。这种血红蛋白有许多性质与HbA不同。它具有明显的抗碱性和抗酸性,人们可以利用这种特性进行测定HbF的含量,检测红细胞中HbF的“酸洗脱”技术就是根据此特性建立的。组成HbF的γ链中含有4个异亮氨酸(Ile),而HbA 和HbA2中不含有这种氨基酸。抗原性方面,HbF也比HbA强。HbF的电泳行为和层析性质也与HbA有些不同,在碱性缓冲液中,其电泳速度稍慢于HbA,用DEAE-纤维素柱层析分离脐带血时,HbF稍后于HbA被洗脱下来。这些性质已广泛用于HbF的分离和研究。HbF的另一个重要性质在于它在生理条件下对氧的亲和性明显高于HbA,这就意味着任何一定的氧分压情况下,HbA取向放出氧到HbF上,将氧从母体转移至胎儿的环境中(下图),显然,这对处在低氧紧张期的胎儿是有益的。
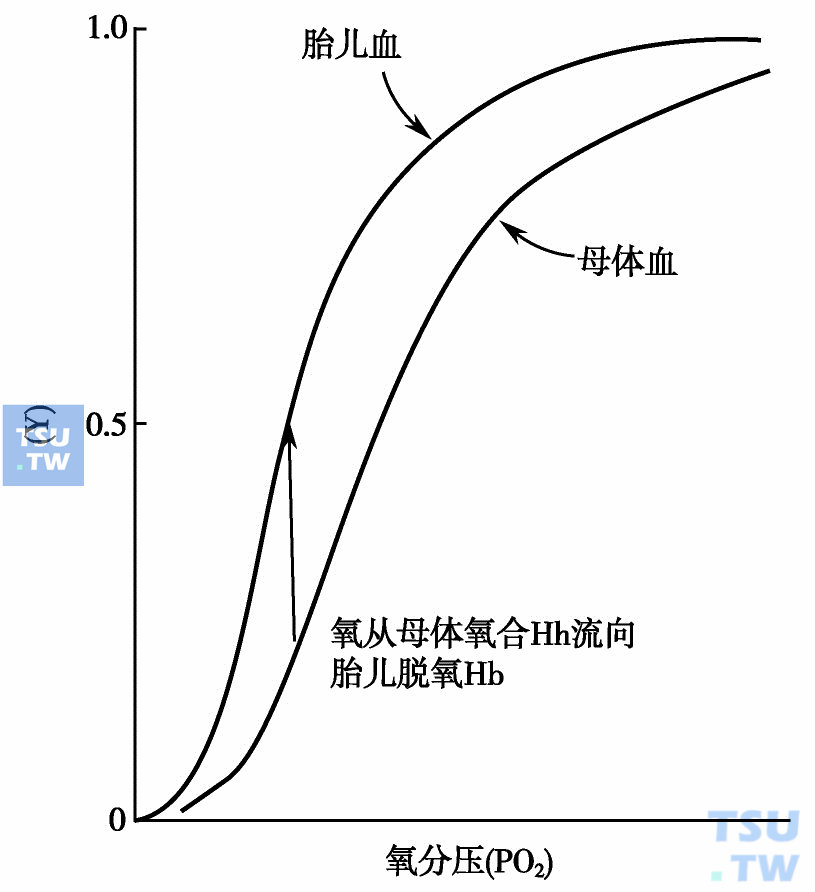
胎儿血和母体血氧解离曲线图
在成人血液中HbF约占总血红蛋白的1%,用酸洗脱或荧光免疫技术分析红细胞中HbF的分布,发现HbF在各红细胞中不是均匀分布的,并非所有红细胞中γ珠蛋白含量都占β珠蛋白含量的1%左右,事实上,多数红细胞都不含HbF,而有些红细胞中HbF占Hb总量的10%~30%,这一类红细胞称为F细胞。因此在成人血液中至少有两种红细胞,一种是F细胞,它既合成β珠蛋白,又合成γ珠蛋白;另一种红细胞则合成β珠蛋白。1986年Schroder和Huisman用聚丙烯酰胺凝胶酸性电泳方法进行珠蛋白肽链分析,发现人类胎儿HbF中的γ珠蛋白链存在两种类型,一种在肽链的136位为甘氨酸(Gly),另一种在肽链的136位为丙氨酸(Ala),分别称为Gγ和Aγ链。它们是Gγ和Aγ两个不等位基因的表达产物。大量新生儿脐带血分析结果表明,Gγ和Aγ链的平均值分别为70%和30%(即Gγ/ Aγ比值为7:3),在整个胎儿时期,Gγ/Aγ这个比值较稳定。在出生后头几个月Gγ/Aγ比值由7:3变为6:4。出生后6个月后,这个比值变为成人型的4:6。现在查明这两种γ链存在于所有人群中,也就是说在正常人血液中含有两种HbF,一种由一对α链和一对Gγ链组成(α2Gγ 2),另一种由一对α链和一对Aγ链组成(α2Aγ2)。并发现有些新生儿Gγ/Aγ比值明显高于7:3。基因图谱分析发现这些新生儿常有γ珠蛋白基因的异常,发生γ基因重排。
另外还发现一些γ珠蛋白变异体。Hb Sardinin是最先发现的一种γ链变异体,它的结构特点是γ链的136位为丙氨酸(Ala),75位异亮氨酸(Ile)为苏氨酸(Thr)取代,用Aγ75Ile→Thr表示,简称AγT链,实际上是AγI链变异体。用聚丙烯酰胺凝胶电泳(PAGE)方法一般不能将AγT和AγI分开。1984 年Shelton用反相高效液相层析(RP-HPLC)方法成功地将AγT和AγI链分开(下图)。Huisman等人改进和应用上述分析方法分析了大量不同人种和民族的脐带血标本,发现Hb Sardinin是一种常见的Aγ链变异体,在不同人种、民族或人群中发生率有差别,具有种族和民族特异性,如在意大利人中AγT链发生率为38%,南斯拉夫人中为46%,保加利亚人中为39%,印度人中为35%,日本人中为31%,澳大利亚人中为12%。中国医学科学院梁植权教授实验室检测了我国汉族,西藏藏族,贵州苗族,广西壮族、侗族,辽宁满族,新疆维吾尔族、回族和哈萨克族等民族新生儿HbF中AγT链发生率,与世界其他不同人群(或民族)相比较,发现我国汉族新生儿中,这种变异体发生率为7%~10%;广西壮族和辽宁满族中未发现存在有AγT链;贵州苗族约有12%新生儿中含有AγT链。值得注意的是西藏藏族和新疆哈萨克族新生儿血液HbF中含有AγT链分别高达30%和38%。
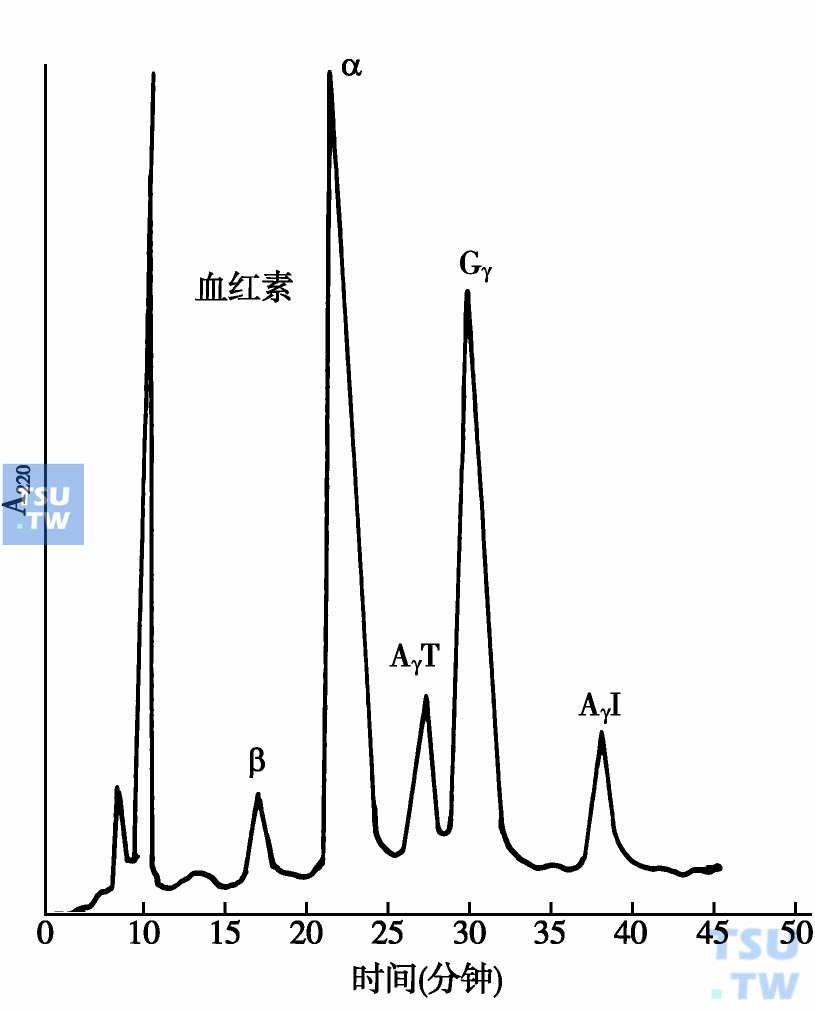
人脐带血中各种珠蛋白链的HPLC分离图
近来与临床直接相关的一项研究是糖基化血红蛋白(血红蛋白合成过程中翻译后修饰产物)的工作。用硼酸盐缓冲液进行凝胶电泳时,我们常常可见到在HbA前面(阳极侧)出现与HbA界限不清、不易分开的次要的扩散成分,Kunke等人曾将这个阳极侧扩散分成称为HbA3,后改名为HbAI。正常的HbA3占HbA的不到10%。柱层析方法可将HbAI分为若干次要成分,分别命名为HbAIa,AIb,AIc。其中HbAIc占4%~6%,HbAIa、HbAIb各占HbA 的1%~2%。研究证实这几种血红蛋白都含有糖类成分。HbAIc与HbA的区别在于其β-链的N-末端缬氨酸残基上结合一个糖类成分,所以人们称其为“糖基化血红蛋白(glycosylated hemoglobin)”。最近分析认为所结合的糖是葡萄糖。HbAIa和HbAIb的结构还不够清楚。由于在红细胞整个生存时期都在进行着HbA缓慢转变形成HbAI(HbA→HbAI),所以在老年红细胞中HbAI要多一些,在幼年红细胞中就要少一些。这个反应是非酶促的,反应速率取决于有效地血糖浓度,后者浓度增高时HbAIc百分比加大。
早在20世纪60年代,人们就注意到在糖尿病患者HbAIc的百分率比非糖尿病的人高。但是关于HbAIc与糖尿病的关系,情况比较复杂。现有资料表明,这种血红蛋白的水平与糖尿病的病程长短并无明显相关,但与糖尿病的严重程度有一定关系。当新糖尿病患者第一次用胰岛素治疗,需用几周后HbAIc百分比才能下降,因此HbAIc百分比的测定反映6~8周前血糖结合值,为此对于评价糖尿病长期控制提供一个有价值的指标。
胚胎血红蛋白,在早期胚胎中ζ-珠蛋白链和ε-珠蛋白链首先合成,其后α-珠蛋白链和γ-珠蛋白链开始合成,于是由这些肽链组成三种胚胎期血红蛋白。最早出现的血红蛋白是由两链条ζ-珠蛋白链和两条ε-珠蛋白组成的HbGower-Ⅰ(ζ2ε2)和由两条α-珠蛋白链和两条ε-珠蛋白组成的HbGower-Ⅱ(α2ε2),随后出现了由两条ζ-珠蛋白链和两条γ-珠蛋白链组成的HbPortland(ζ2γ2)。虽然HbPortland在胚胎期含量并不高,但在正常的脐带血中仍可检测到微量的HbPortland,它可能是由于ζ-珠蛋白链在整个发育过程中持续存在时间比ε-珠蛋白链长一些所致。最近发现,在α-地中海贫血新生儿血液中往往可检测到较高含量的ζ-珠蛋白,因此临床学家正把脐带血中ζ-珠蛋白链的含量作为初步诊断α-地中海贫血的标志之一。ε-珠蛋白和ζ-珠蛋白基因的核酸序列测定已完成,它们的表达产物ε-珠蛋白链和ζ-珠蛋白链分别与β-珠蛋白链和α-珠蛋白链一样,分别由146个和141个氨基酸残基组成,其序列见表:
人体珠蛋白链的一级结构