
趋化因子的发现
早就有人指出入外周血中的单核细胞可能产生一种对中性粒细胞具有趋化作用的因子,但一直未找到。同时也注意到一些细胞因子如白细胞介素-1(interleukin-1,IL-1)、肿瘤坏死因子(tumor necrosis factor, TNF)等对中性粒细胞并无直接的趋化作用,但在体内却可引起中性粒细胞浸润,这一现象始终没有得到满意的解释。1987年,在美国国立癌症研究所(NCI)工作的松岛、吉村从脂多糖(LPS)刺激的人外周血单核细胞培养上清中分离和克隆出了一种对中性粒细胞具有趋化、激活作用的因子,命名为白细胞介素-8(interleukin-8,IL-8, CXCL8)。1989年,他们又分离和克隆了另一种对单核细胞具有趋化、激活作用的因子,命名为单核细胞趋化与激活因子(monocyte chemotactic and activating factor, MCAF)或单核细胞趋化蛋白-1(monocyte chemoattractant protein-1, MCP-1, CCL2),由此拉开了趋化因子研究的序幕。鉴于在IL-1、TNF的刺激下多种细胞均可产生CXCL8,人们终于明白原来IL-1、TNF在体内是通过趋化因子介导引起中性粒细胞浸润的。
趋化因子及其受体的发现是人类征服疾病过程中的重要里程碑,并由此诞生了一门崭新的学科——趋化因子学(chemokinology)。目前该领域的多学科研究已经和正在揭开不少趋化因子及其受体神秘的面纱,以趋化因子及其受体为靶点的新药呼之欲出。趋化因子及其受体的发现不仅解开了白细胞浸润的类型特异性之谜,也为许多疾病的高效和特异性治疗带来了新的希望。
趋化因子的系统命名
趋化因子(chemokine)是由chemotactic cytokine缩写而来,亦称趋化性细胞因子。趋化因子超家族是可特异性募集和激活白细胞的一大类小相对分子质量(8 000~10 000)蛋白质(含70~100个氨基酸残基),依其分子结构中氨基端半胱氨酸(cysteine,C)残基的排列顺序,至少可分为C、CC、CXC和CX3C4个家族。其中C族的代表为XCL1,CC族的代表为CCL2,CXC族的代表为CXCL8,CX3C族的代表为CX3CL1。一般认为,CXC族趋化因子主要作用于中性粒细胞,而C、CC和CX3C族趋化因子主要作用于单核-巨噬细胞及淋巴细胞。趋化因子为碱性蛋白质,可通过与血管内皮细胞表面蛋白聚糖中的含糖醛酸、糖胺聚糖(GAG)相互作用,在局部形成一个引导白细胞定向运动必不可少的浓度梯度。
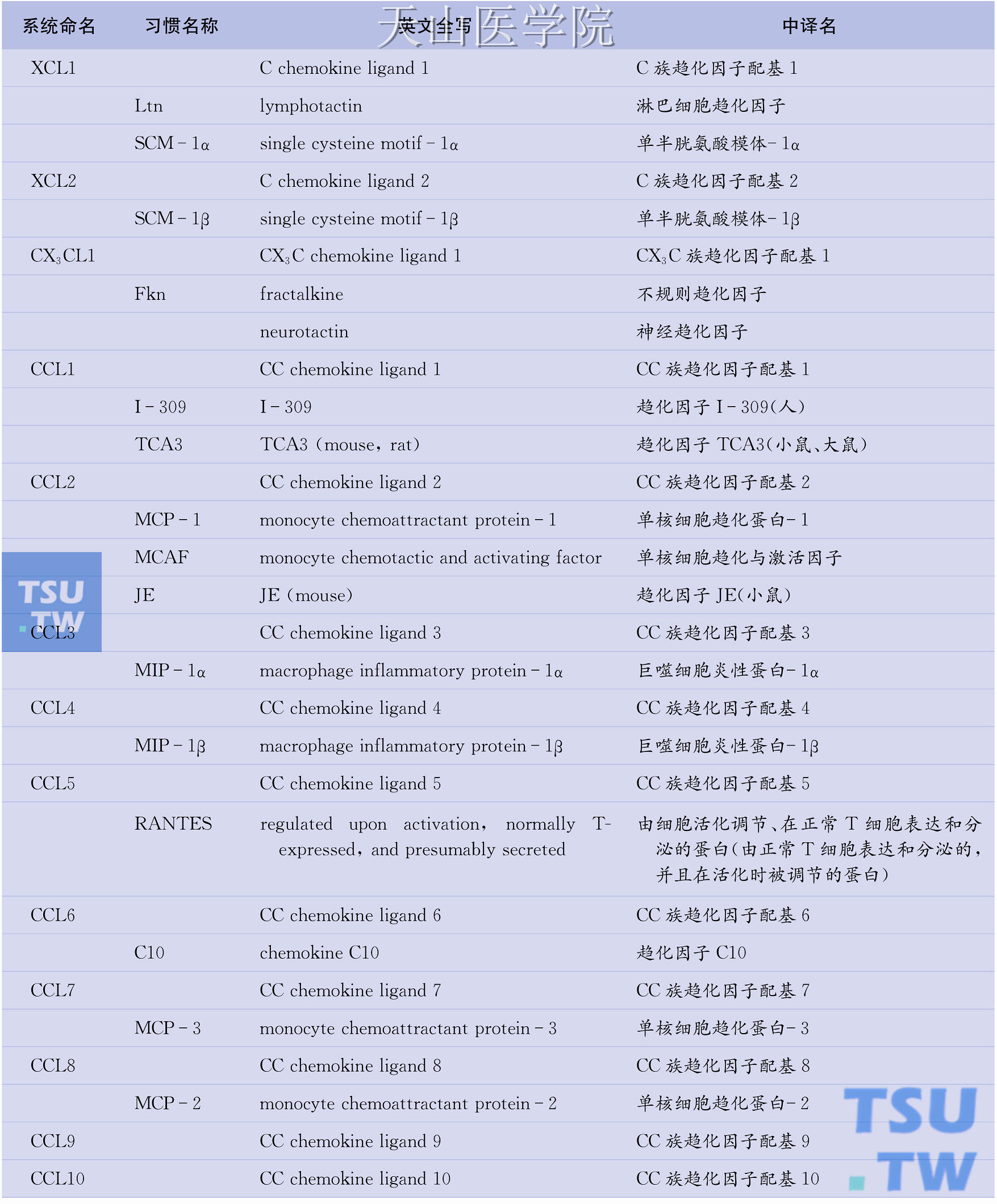
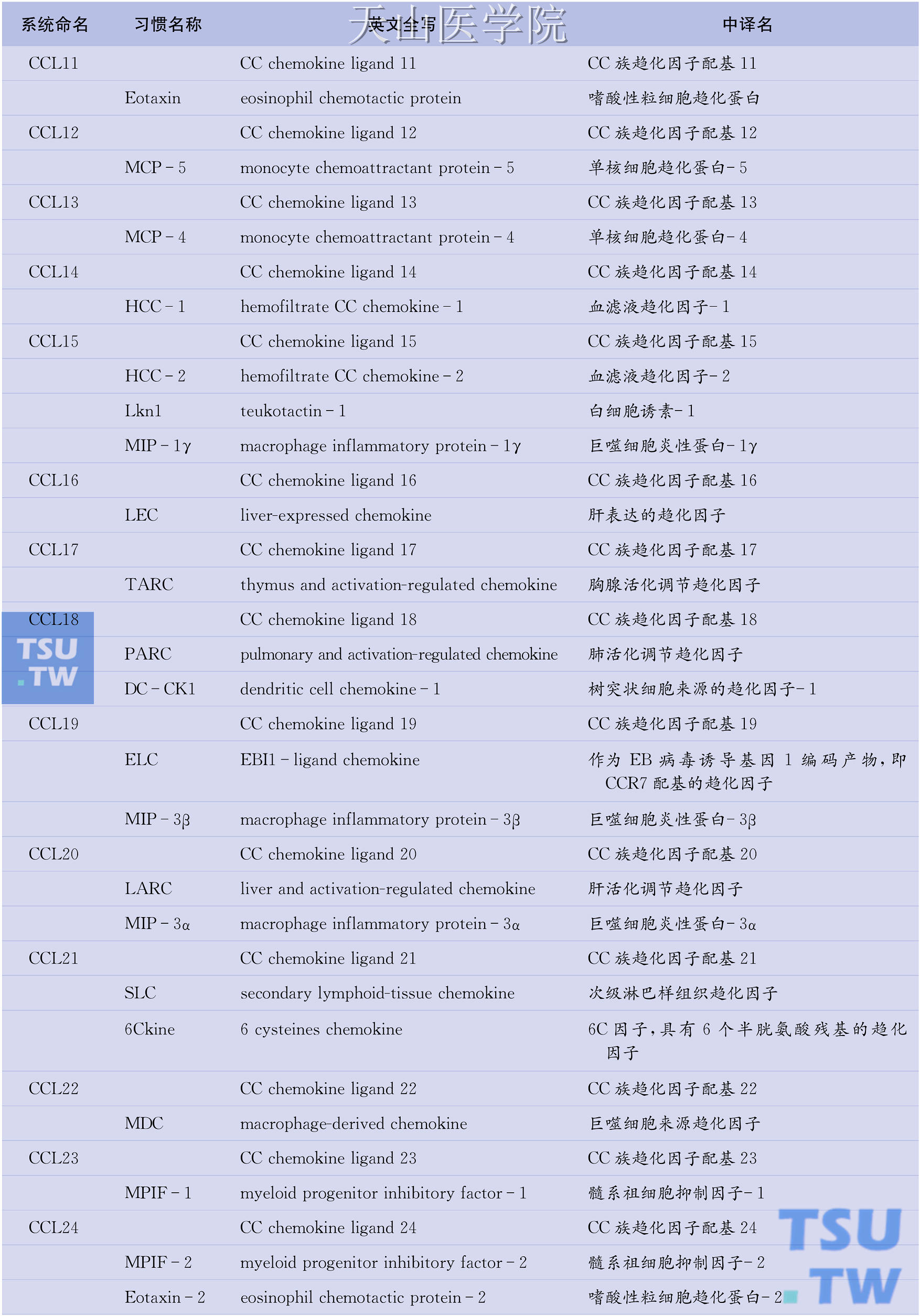
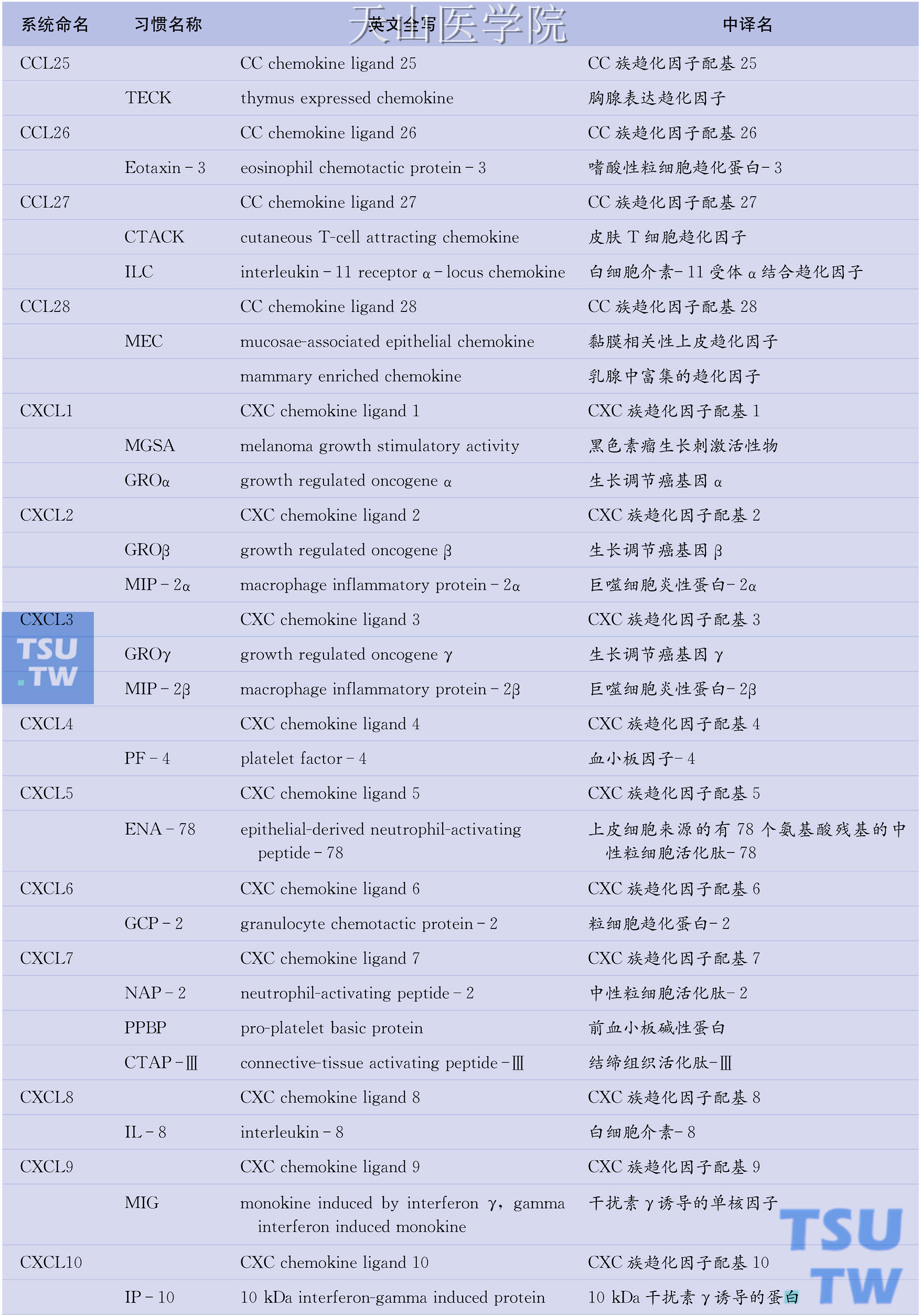
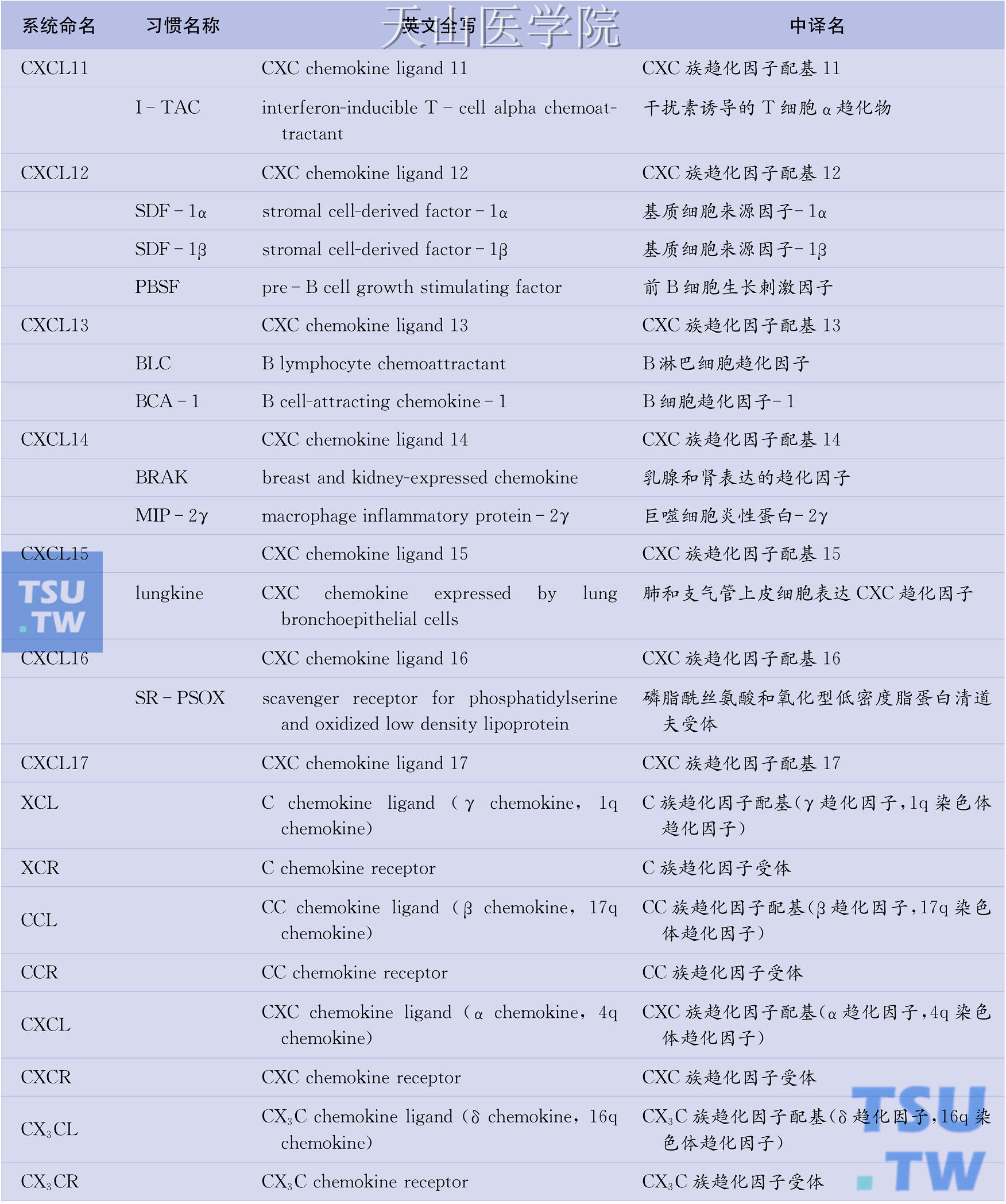
目前已知的人类趋化因子逾50种,趋化因子受体也有20种之多。问题是趋化因子的命名较为混乱,一物多名现象普遍存在。例如,作为CC族趋化因子代表的单核细胞趋化蛋白-1(MCP-1)也称单核细胞趋化与激活因子(MCAF)、淋巴细胞衍生趋化因子(LDCF)、神经胶质瘤衍生趋化因子(GDCF)、肿瘤衍生趋化因子(TDCF)或平滑肌细胞衍生趋化因子(SMC-CF),而在小鼠则称为JE基因产物。因此,系统命名趋化因子势在必行。所幸的是,已有学者提出了对趋化因子进行系统命名的方案(表6-1),该方案吸收了现行趋化因子受体命名法(如CXCL8的受体CXCR1、MCP-1的受体CCR2)一目了然的优点,采用分子结构特征(C、CC、CXC、CX3C)+配基(ligand, L)+序号(1,2,3…)的方法对各种趋化因子重新命名。如IL-8属于CXC族,其系统名定为CXCL8。同样,属于CC族的MCP-1的新名为CCL2;属于C族淋巴细胞趋化因子的新名为XCL1;属于CX3C族不规则趋化因子的新名为CX3CL1。需要特别注意的是,除CXCL2、XCL1、CX3CL1、CCL2等个别趋化因子外,系统名中的序号与趋化因子受体名中的序号并不对应。
表6-1 趋化因子的系统命名
趋化因子的功能
趋化因子是白细胞特异性浸润所必需的
经典的白细胞趋化剂如细菌产物甲酰三肽(fMLP)、补体片段C5a、脂类介质血小板激活因子(PAF)、白细胞三烯-4(LTB-4)等虽可引起白细胞浸润,但缺乏白细胞类型特异性。此外,除去补体片段C5a并不影响白细胞浸润,可见它们不是白细胞浸润所必需的。
趋化因子的发现解开了白细胞浸润的类型特异性之谜。不同趋化因子的白细胞趋化特异性是由各种白细胞膜上趋化因子受体的表达水平所决定的。如中性粒细胞表达CXCR1、CXCR2;单核-巨噬细胞表达CCR1、CCR2、CCR5;嗜酸性粒细胞表达CCR3;T细胞上的趋化因子受体分布相当复杂,因不同时相和细胞亚型而异,包括CXCR3~5、CCR1~7、CCR9、XCR1、CX3CR1等。
白细胞由血流外渗到组织至少包括滚动、黏附和渗出3个步骤:①滚动。通过白细胞表面的选择素(selectin)受体与血管内皮细胞表面的选择素相互作用,白细胞沿着血管内壁滚动。②黏附。呈碱性的趋化因子可结合于血管内皮细胞表面含硫酸类肝素的蛋白聚糖和细胞外基质分子上,作为过往白细胞的信号。当滚动中的具有相应趋化因子受体的白细胞与此信号趋化因子结合后即被活化,引起白细胞表面作为黏附分子受体的整联蛋白(如VLA-4/整联蛋白41、LFA-1/整联蛋白L2)发生立体变构而活化,从而与血管内皮细胞表面的黏附分子配体[如血管细胞黏附分子-1(VCAM-1)、细胞间黏附分子-1(ICAM-1)]形成较牢固的结合。③渗出。被捕获的白细胞沿着由游离趋化因子构成的浓度梯度透过血管内皮进入组织间隙。显而易见,在此过程中趋化因子起着关键性的作用。
趋化因子受体CCR5基因缺损者可抵抗人类免疫缺陷病毒感染
1996年,有关人类免疫缺陷病毒(human immunodeficiency virus,HIV)利用某些趋化因子受体作为第二受体而侵入并感染CD4+T细胞及巨噬细胞的发现引起了极大的轰动,也使趋化因子研究揭开了新的一页。
以艾滋病患者血中CD4+Th细胞极度减少为线索,现已证实CD4是HIV侵入细胞的第一受体。然而,导入人CD4基因的动物细胞并不受HIV感染,提示在人的易感细胞上可能存在着CD4之外的特异性分子。1996年,Berger等首先报道融合素(fusin,即CXCL12的受体CXCR4)是T细胞向性 HIV的协同受体。几乎与此同时,其他学者的研究表明CCR5(CCL3、CCL4和CCL5的受体)是巨噬细胞向性HIV的协同受体。
HIV以趋化因子受体为其第二受体的重大发现使人们理解了一些趋化因子如CXCL12、CCL3、CCL4和CCL5缘何具有抗HIV作用。令人惊异的是,CCR5基因32bp缺损的个体可以抵抗HIV感染。在白种人中持有这种变异者约占10%,其中纯合子为1%,杂合子为18%。携带CCR5基因变异的纯合子对HIV感染具有高度抵抗性,在杂合子其发病可延迟2~4年。
趋化因子参与炎症、免疫、造血、血管生成及器官发育
趋化因子主要与炎症有关,故有“炎症性趋化因子”一说。但趋化因子不仅参与炎症反应,还涉及免疫、造血、血管生成及器官发育等,因此将这些作用广泛的多能性因子称作趋化因子甚至炎症性趋化因子实非所宜。
近年来发现的CCL17~21等趋化因子与淋巴组织的形成、维持免疫应答密切相关,因而提出“免疫系趋化因子”的概念。在CXCL12及其受体CXCR4基因敲除小鼠,可见骨髓造血障碍、胃肠血管异常和心脏膜性室间隔缺损。CCL5在胚胎期的肾小球系膜区和近曲小管有较强的持续性表达,故认为这种CC族趋化因子可能与人的肾脏发育有关。在CXC族趋化因子中含谷氨酸-亮氨酸-精氨酸基序(glutamate-leucine-arginine motif, ELR基序)者如CXCL1-3、CXCL5-8具有促进血管生成作用,而无ELR基序者如CXCL4、CXCL9~14具有抑制血管生成的作用(CXCL12例外)。据报道CXCL10可抑制非小细胞肺癌的血管生成和转移,而抑制CXCL8也可抑制肺癌的血管生成。此外,在CCL2受体CCR2缺损的小鼠,动脉硬化病灶中巨噬细胞浸润及动脉硬化病变的程度均明显减轻。CCL17、CCL28等趋化因子还具有直接的抗微生物活性。
