
遗传物质:DNA
脱氧核糖核酸(DNA)是生物的遗传物质。后来的研究证明,绝大多数生物体的遗传物质是DNA,有些病毒的遗传物质是核糖核酸(RNA)。
1953年.Watson和Crick创立了DNA双螺旋结构模型,发现了DNA中四种碱基的比例关系为A/T=G/C=1;阐明了DNA分子的结构特征,而且提出了DNA作为执行生物遗传功能的分子,从亲代到子代的DNA复制(replication)过程中,遗传信息的传递方式及高度保守性。DNA双螺旋结构模型的确立为遗传学进入分子水平奠定了基础,是现代分子生物学的里程碑。
Crick于1958年提出的分子遗传中心法则(central dogma)揭示了核酸与蛋白质间的内在关系,以及RNA作为遗传信息传递者的生物学功能。并指出了信息在复制、传递及表达过程中的一般规律,即DNA-RNA-蛋白质。遗传信息以核苷酸顺序的形式贮存在DNA分子中,它们以功能单位在染色体上占据一定的位置构成基因(gene)。
1975年,Sanger发明DNA测序方法。人有46条染色体(22对常染色体+性染色体X,Y),单倍体基因组有3x109碱基对,测序已经完成。
基因
基因是具有遗传效应的DNA分子片段,在染色体上呈线性排列。基因不仅可以通过复制把遗传信息传递给下一代,还可以使遗传信息得到表达,也就是使遗传信息以一定方式反映到蛋白质的分子结构上,从而使后代表现出与亲代相似的性状。
“基因”一词,是由“GENE”音译而来的。20世纪30年代,由于证明了基因是以直线的形式排列在染色体上,因此人们认为基因是染色体上的遗传单位。随着分子遗传学的发展,Watson和Crick在1953年提出DNA的双螺旋结构以后,人们普遍认为基因是DNA的片段,确定了基因的化学本质。大多数生物的基因由DNA组成,而DNA则是染色体的主要化学成分。大多数真核生物细胞内的DNA是由双股多核苷酸单链结合而成。每股DNA链又由许多个单核苷酸借磷酸二酯键互相连接而成;而两股单链之间则是依靠两者的碱基成分按互补规律分别配对结合,即腺嘌呤(A)与胸腺嘧啶(T)以两个氢键配对,鸟嘌呤(G)与胞嘧啶(C)以三个氢键配对,形成双螺旋结构,故称为DNA双螺旋。20世纪60年代,Benz提出了基因的内部具有一定的结构,可以区分为突变子、互换子和顺反子三个不同的单位。DNA分子上的—个碱基变化可以引起基因突变,因此可以看成是一个突变子;两个碱基之间可以发生互换,可以看成是一个互换子;一个顺反子是具有特定功能的一段核菅酸序列,作为功能单位的基因应该是顺反子。因此从分子水平来看,基因就是DNA分子上的一个片段,经过转录和转译能合成一条完整的多肽链。可是,通过近来的研究认为这个结论并不全面,因为有的基因在转录出RNA后,不再翻译成蛋白质。另外,还有一类基因,如操纵基因,它们既没有转录作用,又没有转译产物,仅仅起着控制和操纵基因活动的作用。特别是近年来还发现DNA分子上有相当一部分片段,只是某些碱基的简单重复。这类不含有遗传信息的碱基片段,在真核细胞生物中数量可以很大,甚至达到50%以上。关于DNA分子中这些重复碱基片段的作用,目前还不十分了解。有人推测可能有调节某些基因活动和稳定染色体结构的作用,其真正的功能尚待研究。由此,目前认为应把基因看作是DNA分子上具有特定功能的(或具有一定遗传效应的)核苷酸序列。
基因的一般结构
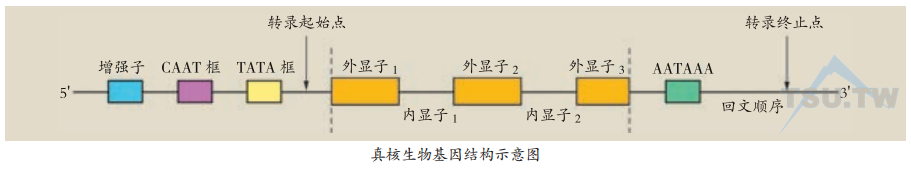
高等真核生物的绝大多数基因是断裂的。一个断裂基因含有若干段编码序列,这些可以编码的序列称为外显子。在两个外显子之间被一段不编码的间隔序列隔开,这些间隔序列称为内含子。每个断裂基因在第一个和最后一个外显子的外侧各有一段非编码区,为侧翼序列。在侧翼序列上有一系列调控序列,主要包括启动子(promoter)、增强子(enhancer)和终止序列。
启动子主要包括以下四种元件:
- 在转录起始点上游约20~30bp处有TATA框(TATA box)。TATA框是一个短的核苷酸序列,其碱基序列一般为TATAATAAT。TATA框是RNA聚合酶的一个重要的识别位点,它能够使酶准确地识别转录的起始点并开始转录,对核心启动子的定位有决定性作用,决定转录起始的位置。
- 在转录起始点上游约70~80 bp处有CAAT框(CAAT box)。CAAT框是启动子中另一个短的核菅酸序列,其碱基序列一般为GGCTCAATCT。CAAT框正反向排列均起作用,是RNA聚合酶的另一个结合点,在决定启动子的效率方面发挥重要作用,但似乎对启动子的特异性没有直接作用,它的存在增强了启动子的强度。
- 在转录起始点上游约90 bp处有GC框(GC box)。其碱基序列一般为GGGCGG,在启动子中经常以多拷贝存在,它们以正反两个方向排列,也是相对常见的启动子序列。
- 八聚体元件。
启动子按匹配原则进行组织,多种不同元件同时存在可以促进启动子发挥作用,但没有一个元件是所有启动子都需要的,在任何一个启动子中发现的这些元件在数目及位置排列方向上都不相同。核心启动子元件和上游启动子元件有不同的功能。核心启动子元件(如TATA框和起始点)单独起作用时只能确定转录起始位点和产生基础水平的转录。而上游启动子元件(如GC框或CAAT框)与相应的蛋白因子结合能提高或改变转录效率。不同基因具有不同的上游启动子元件组成,其位置也不相同,就使得不同的基因表达分别有不同的调控。
增强子是一种能够提高转录效率的顺式调控元件。最早是在SV40病毒中发现长约200 bp的一段DNA,可使旁侧的基因转录效率提高100倍,其后在多种真核生物,甚至在原核生物中都发现了增强子。增强子通常占100~200 bp长度,也和启动子一样由若干组件构成,其基本核心组件常为8~12 bp,可以单拷贝或多拷贝串连形式存在。
增强子的位置不固定,可以位于启动子的上游或下游,而且能在正反两个方向发挥作用。增强子要有启动子才能发挥作用,没有启动子存在,增强子不能表现活性。但增强子对启动子没有严格的专一性,同一增强子可以影响不同类型启动子的转录,能影响任何位于它附近的启动子,也可以远距离起作用。增强子的作用机制虽然还不明确,但与其他顺式调控元件一样,必须与特定的蛋白质因子结合后才能发挥增强转录的作用。增强子一般具有组织或细胞特异性。此外,所有的增强子中均有一段由交替的嘧啶-嘌呤残基组成的DNA,这种DNA极易形成Z-DNA型;故有人认为在形成一小段Z-DNA后,增强子才有功能。
终止序列在终止密码的下游,真核生物转录终止的机制尚知之甚少。RNA聚合酶I转录出rRNA前体3’末端后,继续向下游转录超过1 000个碱基,此处有一个18 bp的终止序列,在辅助因子的辅助下转录终止。内切核酸酶再切割产生rRNA前体的3’末端。
RNA聚合酶Ⅲ转录模板的下游存在一个终止子,该终止子是位于GC丰富序列之中的TTTT(编码链)。这是所有真核生物RNA聚合酶Ⅲ转录的终止信号。这种序列的特征是高度保守,从酵母到人都很相似,任何改变这种序列特征的突变都将导致转录终止位置的改变。RNA聚合酶Ⅲ有内源性的转录终止功能,能识别终止子,使转录终止。终止位点一般在第二个T,但也可能在第三或第四个T,因此RNA聚合酶Ⅲ催化合成的RNA前体3’端有U序列。真核生物mRNA3’端有多聚腺苷酸(polyA)尾巴,这是转录后才加进去的,因为在模板链上没有相应的polyT序列。在结构基因最后一个外显子的3’端,常有一组共同序列AATAAA,其下游还有相当多的GT序列。这些序列称为转录修饰点。当RNA聚合酶Ⅱ转录出AAUAAA后,聚腺苷酸特异因子(CPSF)能识别它并与它结合。在CPSF以及切割活化因子(CstF)等因子的指导下,特异的内切核酸酶在AAUAAA下游11-30核苷酸处切断RNA链,mRNA前体(hnRNA)的转录即告终止。
基因突变(mutation)及基因多态性(polymorphism)
基因突变是指由于DNA碱基对的置换、增添或缺失而引起的基因结构的变化,亦称点突变。在自然条件下发生的突变叫自发突变,由人工利用物理因素或化学药剂诱发的突变叫诱发突变。基因突变是生物变异的主要原因,是生物进化的主要因素。人工诱变是产生生物新品种的重要方法。
根据基因结构的改变方式,基因突变可分为碱基置换突变和移码突变两种类型。
碱基置换突变:由一个错误的碱基对替代一个正确的碱基对的突变叫碱基置换突变。例如在DNA分子中的GC碱基对由CG、AT或TA所代替,AT碱基对由TA、GC或CG所代替。碱基替换过程只改变被替换碱基的那个密码子,也就是说每一次碱基替换只改变一个密码子,不会涉及其他的密码子。引起碱基置换突变的原因和途径有两个:一是碱基类似物的掺入,例如DNA的一部分胸腺嘧啶可被5-溴尿嘧啶(BU)所取代,从而导致AT碱基对变成GC碱基对,或者GC碱基对变成AT碱基对。二是某些化学物质如亚硝酸、亚硝基胍、硫酸二乙酯和氮芥等,以及紫外线照射,也能引起碱基置换突变。
移码突变:基因中插入或者缺失—个或几个碱基对,会使DNA的阅读框架(读码框)发生改变,导致插入或缺失部位之后的所有密码子都跟着发生变化,结果产生一种异常的多肽链。移码突变诱发的原因是一些像吖啶类染料分子能插入DNA分子,使DNA复制时发生差错,导致移码突变。
根据遗传信息的改变方式,基因突变又可以分为同义突变(same sense mutation)、错义突变(missense mutation)和无义突变(nonsense mutation)三种类型。
同义突变:有时DNA的—个碱基对的改变并不会影响它所编码的蛋白质的氨基酸序列,这是因为改变后的密码子和改变前的密码子是兼并密码子,它们编码同一种氨基酸,这种基因突变称为同义突变。
错义突变:由于一对或几对碱基对的改变而使决定某一氨基酸的密码子变为决定另一种氨基酸的密码子的基因突变叫错义突变。这种基因突变有可能使它所编码的蛋白质部分或完全失活,例如人血红蛋白B链的基因如果将决定第6位氨基酸(谷氨酸)的密码子由cTT变为CAT,就会使它合成出的β链多肽的第6位氨基酸由谷氨酸变为缬氨酸,从而引起镰刀状细胞贫血病。
无义突变:由于一对或几对碱基对的改变而使决定某一氨基酸的密码子变成—个终止密码子的基因突变叫无义突变。
基因突变作为生物变异的—个重要来源,它具有以下主要特点:
第一,基因突变在生物界中是普遍存在的。无论是低等生物,还是高等的动植物以及人,都可能发生基因突变。
第二,基因突变是随机发生的。它可以发生在生物个体发育的任何时期和生物体的任何细胞。一般来说,在生物个体发育的过程中,基因突变发生的时期越迟,生物体表现突变的部分就越少。基因突变可以发生在体细胞中,也可以发生在生殖细胞中。发生在生殖细胞中的突变,可以通过受精作用直接传递给后代。发生在体细胞中的突变,一般是不能传递给后代的。
第三,在自然状态下,基因突变的频率是很低的。在高等生物中,大约10万个到l亿个生殖细胞中,才会有一个生殖细胞发生基因突变,突变率是1/(105~108)。不同生物的基因突变率是不同的。例如,细菌和噬茵体等微生物的突变率比高等动植物的要低。同一种生物的不同基因,突变率也不相同。
第四,大多数基因突变对生物体是有害的。由于任何一种生物都是长期进化过程的产物,它们与环境条件已经取得了高度的协调。如果发生基因突变,就有可能破坏这种协调关系。因此,基因突变对于生物的生存往往是有害的。例如,绝大多数的人类遗传病,就是由基因突变造成的,这些病对人类健康构成了严重威胁。但也有少数基因突变是有利的。例如,植物的抗病性突变、耐旱性突变,微生物的抗药性突变等,都是有利于生物生存的。
第五,基因突变是不定向的。—个基因可以向不同的方向发生突变,产生一个以上的等位基因。
基因多态性是指处于随机婚配的一个群体中,同一基因位点可存在两种以上的基因型。在人群中,个体间基因的核菅酸序列存在着差异性称为基因(DNA)的多态性。这种多态性可以分为两类,即DNA位点多态性(site polymorphism)和长度多态性(length polymorphism)。基因的多态性显示了遗传背景的多样性和复杂性。它可能是人类在进化过程中抵御不良环境因素的一种适应性表现,对维持种群的生存与延续具有重要的生物学意义。同一基因位点的不同基因型叫等位基因(allele),等位基因的分布频率在随机婚配群体中符合Hardy-Weinberg平衡定律。
所谓Hardy-Weinberg平衡定律,是指在一个不发生突变、迁移和选择的无限大的相互交配的群体中,基因频率和基因型频率将逐代保持不变。Hardy-Weinberg平衡定律可分为三个部分:第一部分是假设在一个无穷大的随机交配的群体中,没有进化的压力(突变、迁移和自然选择);第二部分是基因频率逐代不变;第三部分是随机交配一代以后基因型频率将保持平衡:p2表示AA的基因型的频率,2pq表示Aa基因型的频率,q2表示aa基因型的频率。其中p是A基因的频率;q是a基因的频率。基因型频率之和应等于1,即p2+2pq+q2=1。这个定律简而言之:在没有进化影响下当基因一代一代传递时,群体的基因频率和基因型频率将保持不变。
位点多态性:是由于等位基因之间在特定的位点上DNA序列存在差异,也就是基因组中散在的碱基的不同,包括点突变(转换和颠换),单个碱基的置换、缺失和插入。单个碱基的变异又称为单核苷酸多态性(single nucleotide polymorphism,SNP),SNP通常是一种二等位基因(biallelic)或二态的变异。据估计,单碱基变异的频率在1/1 000~2/1 000。SNP在基因组中数量巨大,分布频密,检测易于自动化和批量化,被认为是新一代的遗传标记。突变可以看作是SNP的一种特殊形式,一般认为变异位点在人群中的频率大于1%即为SNP,反之则为突变。
长度多态性:是基因组内的重复序列。可变数目串联重复序列(variable number of tandem repeats,VN-TRs)是由于相同的重复序列重复次数不同所致,它决定了小卫星DNA( minisatellite)长度的多态性。小卫星是由15~65 bp的基本单位串联而成,重复单位长度通常不超过20bp,重复次数在人群中是高度变异的。微卫星DNA (microsatellite),基本序列只有1-8bp,如(TA)n及(CGG)n等,通常重复10-60次,因此又叫短串联重复序列(short tandem repeats,STR)。长度多态性按照盂德尔方式遗传,STR在基因定位、DNA指纹分析、遗传病的分析和诊断中被广泛应用。
基因多态性的医学意义:人类基因多态性在阐明人体对疾病、毒物的易感性与耐受性,疾病临床表现的多样性(clinical phenotype diversity),以及对药物治疗的反应性上都起着重要的作用。临床上早期有关基因多态性的研究是从HLA基因开始的,分析基因型在疾病发生易感性方面的作用,如HLA-B27等位基因与强直性脊柱炎发生率的密切关联,可作为诊断的依据。通过基因多态性的研究,可从基因水平揭示人类不同个体间生物活性物质的功能及效应存在着差异的本质。通过对基因多态性与疾病的易感性的联系研究,如p53抑癌基因多态性与肿瘤发生及转移的关系研究,可阐明人体对疾病、毒物和应激的易感性,不仅为临床医学也为预防医学的发展开辟了新的领域。
疾病基因多态性与临床表型多样性的联系已受到重视,如肿瘤等多基因病的临床表型往往多样化,阐明基因型( genotype)与表型(phenotype)之间的联系在认识疾病的发生机制、预测疾病的转归等方面也有重要的作用。药物代谢基因多态性可以影响药物的代谢过程及清除率,从而影响治疗效果。致病基因的多态性使同一疾病不同个体其体内生物活性物质的功能及效应出现差异,导致治疗反应性上悬殊,按照基因多态性的特点用药,将会使临床治疗符合个体化的要求。在疾病基因多态性研究的引导下,临床医生将有可能预测不同的个体在同样的致病条件下会出现什么样的病理反应和临床表现,即临床表型。如高血压的治疗将根据基因多态性的研究选择更具针对性的药物,调整其剂量,而不是不加选择地使用ACEI、钙拮抗剂或交感神经受体阻断剂。合并症的防治也会更个体化,更具针对性。基因多态性的研究对于遗传病具有双重意义:一方面,基因的有害突变,不论是经典的点突变,还是动态突变,其本身就可能是遗传病的病因;另一方面,众多的多态性位点又是很好的遗传标记,可以在遗传病的研究和临床诊断中发挥重要的作用。
- 多态性作为遗传病的病因: 点突变引起的疾病:从镰刀状细胞贫血开始,突变引起各种遗传病的例子愈来愈多,遗传性肿瘤也逐渐被认识。重复序列多态性作为遗传病的病因,如CCG、CTG和CAG这样的三核苷酸重复序列,当其拷贝数过度增高时可以引起强直性肌营养不良等。三核苷酸拷贝数的扩增或突变发生在世代传递过程中,由于拷贝数在世代间的改变,它被称为动态突变。目前动态突变疾病大多是些神经系统的退行性疾病,也有少数肿瘤。动态突变疾病的发现提示序列拷贝数的多态性能够成为遗传病的病因。
- 多态性作为遗传标记:绝大多数DNA多态性并不引起遗传病,但可作为遗传标记来使用。例如:上述提到的各种多态性标记,如STR和SNP位点,都已广泛用于遗传病的连锁分析。利用染色体上位置已知的众多的多态性标记,通过患病家系的连锁分析,可以对遗传病的致病基因定位,并为随后的基因克隆提供依据。此外,在疾病的关联分析和病因学研究方面,通过比较患病群体和正常群体,可以发现两组间多态性位点的特定等位基因频率是否有显著性差别,来研究该位点与该疾病是否相关。使用多态性标记的关联分析既可以提示相关基因存在的位置,也有助于发病机制的阐明。基因多态性还可以用于疾病的分型与治疗,即根据患者疾病多态性的基因型来解释疾病的病因和临床表现。
在预防医学方面,基因多态性的研究涉及的范围广泛,包括基因多态性与病因未知的疾病关系的研究,也包括对已知特定环境因素易感基因的筛选。由于基因多态性有明显的种族差异,因此在基因一环境交互作用模式上,不同的种族之间有可能不同。而基因多态性的研究在职业病医学中则更具有实际的意义。对易感基因和易感性生物标志物的分析,将某些携带敏感基因型的人甄别开来,采取针对性预防措施,提高预防职业性危害工作的效率。对特定的污染物易感人群和耐受人群的基因多态性研究,有助于阐明环境因素的致病机制,也推动了遗传易感性标志物的研究。
