
精子核结构
精子核位于头部中央,体积较小,由一团浓聚的染色质构成,外包核膜。人精子染色质为不规则的纤维颗粒状。在浓密的核染色质中常存在不规则的透亮区,称为核泡(nuclear vesicle),以精子头部的前半多见。核泡是染色质排列发生明显畸变的结果,可影响精子的受精能力。核内含有DNA、RNA、核蛋白、K+、Ca2+、Mg2+、Fe2+、Cu2+等金属离子。其DNA的量和核蛋白的组成都具有独特性,与DNA有关的主要核蛋白是鱼精蛋白。
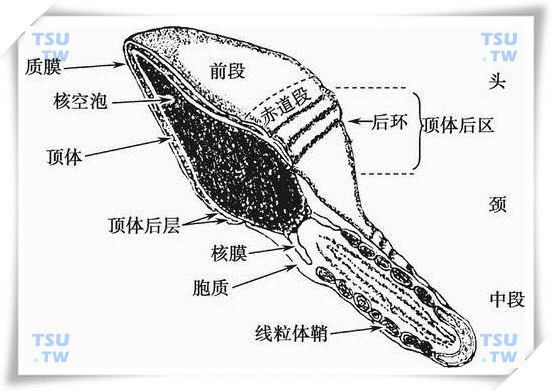
人精子头部超微结构模式图
大部分的核膜无核孔,双层膜间隔7~10nm。在皱褶区却存在较多的核孔,呈六角形;此处的双层膜的间隔与体细胞的核膜一样,为40~60nm。核膜内面有一层蛋白质网,称核板(nuclear lamina),它形成核膜的骨架结构,并可固定染色质。核板由三种密切相关的蛋白质,即核纤肽(lamin)A、B、C构成。
精子核遗传物质
精子发生基因定位于Y染色体q11、23的5、6位点上。用确定Y染色体区间的分子缺失分析方法,已经测得人类精子发生基因位点内的DNA长度约由5亿个核苷酸组成,由Y q11、23延伸至G显带的Y q11、23的中部。
哺乳类动物精子核几乎全部为DNA所充满。在电镜下,小鼠精子核DNA的体积和核体积二者非常接近。由此推测,精子核DNA不可能包装成核小体,因为就核体积而言,如果说包装成核小体的话,那么就需要至少数倍于精子核或更多的空间才行。所以,哺乳类动物精子核DNA就必然具有一特殊的包装形式。已知在精子细胞阶段之前,染色质纤维(25~50nm)连接组蛋白或鱼精蛋白,精子细胞(长形精子细胞)经变态而形成成熟精子后,原来的组蛋白-DNA复合体被高度浓聚的鱼精蛋白-DNA复合体所替代。即鱼精蛋白是精子核DNA的结合蛋白。由于鱼精蛋白与组蛋白间的这种替代机制不清楚,所以,人们寄希望于了解鱼精蛋白是怎样嵌入其间的。有人提出了一个DNA-鱼精蛋白的结合模式,设想鱼精蛋白嵌在DNA小槽的内侧,并计算出鱼精蛋白的几组氨基酸有足够多的正电荷将DNA磷酸基因负电荷完全中和,然后DNA-鱼精蛋白复合体嵌入相邻DNA链的大槽。这样,精子核DNA以肩并肩线性排列的方式包装起来。染色质因鱼精蛋白之间和鱼精蛋白内的共价二硫键而达到稳定。通过这种方式,全部DNA被包装成很小体积。
人们观察到,DNA环绕组蛋白的八聚体形成的螺旋诱导超级螺旋呈双螺旋,这样的超级螺旋之所以称之为反向螺旋,是因为它的方向和Watson-Crick DNA双链螺旋解旋的方向一致。含组蛋白的两栖类动物精子核DNA具有超级螺旋,而含鱼精蛋白的哺乳动物精子核DNA则不发生超级螺旋。由此可知,不发生超级螺旋是哺乳动物精子核DNA浓缩包装机制之一。
电镜下,大鼠和家兔精子核染色质呈板层排列,而这种板层排列平行于核的长轴,也支持精子核DNA呈线性平行排列的模式。人类成熟精子核中组蛋白含15%。鱼精蛋白含量占有极明显优势(85%),故有证据支持这一推测,即DNA分别特异地和组蛋白、鱼精蛋白结合在一起,形成复合体的结构特征。
精子核染色质的平行线性排列是由基质将其组织成袢域,而使DNA形成三维空间构型。精子核DNA的袢域较小,可能体现了它和行将参与的胚胎的相似性。有学者将仓鼠精子核充分解聚后发现,袢域不再附着于核基质,DNA却固着于植入小凹,后者也被称为核小环的特殊结构,这提示每一条染色体至少有一个核小环附着点。虽然说精子无活跃的转录活动,也不复制DNA,但其正常功能对核结构的依赖性却十分重要。
哺乳类动物精子核所呈现的复杂结构,似乎反映了DNA具有功能意义的组织方式。这种组织方式不仅可将致密的严谨顺序的遗传信使导入卵子,而且使这些信使以恰当的生理化学组织方式传递,从而准确无误地进入胚胎。鱼精蛋白将精子DNA包装成致密的小体积,核基质和核小环又将致密的DNA组织成特异的结构。还有研究显示,DNA袢域中有6对基因和仓鼠精子核基质有特殊的拓扑学关系。
尽管说既往一些研究资料提示,精子核DNA袢域以特异的三维空间方式组织在核基质之中。但又不可否认,人类精子核中鱼精蛋白的替代中间型碱性蛋白,中间型碱性蛋白代替组蛋白;这三种蛋白与DNA关系,DNA与核结构联系的分子基础的研究,不是一件轻松的工作。
