
遗传物质的载体——染色体(chromosome)由脱氧核糖核酸(deoxyribonucleic acid,DNA)和蛋白质共同组成。DNA分子由两条反向平行的多核苷酸链围绕同一中心轴相互缠绕形成右手螺旋结构,直径2nm,每环绕1周的高度为3.4nm,含10个碱基对(base pair,bp),两个相邻碱基对之间相距0.34nm。DNA链的骨架位于双螺旋的外侧,由交替出现的亲水脱氧核糖基和磷酸基组成;碱基位于双螺旋内侧,多核苷酸链及碱基间形成氢键配对,碱基相互配对遵守碱基互补原则(A-T配对形成两个氢键,G-C配对形成3个氢键)。染色体中的蛋白质主要包括5种带正电荷,并与DNA结合的碱性组蛋白(histone) H1、H2A、H2B、H3和H4,以及呈酸性的非组蛋白。非组蛋白在参与染色体的构建方面主要起辅佐组蛋白作用。
人类染色体含有四级结构
染色体上的DNA并非以简单的双螺旋线性分子存在,而是采取更紧密的方式置于细胞或细胞核内有限的空间中。在真核细胞(如人类细胞)核内,线性DNA双链围绕在组蛋白的八聚体周围,形成核小体(nucleosome)。前后连接成串珠状,经进一步盘旋、折叠,最终形成染色单体(chromatid);也就是说,染色单体是由核小体的重复单位组成的(下图)。
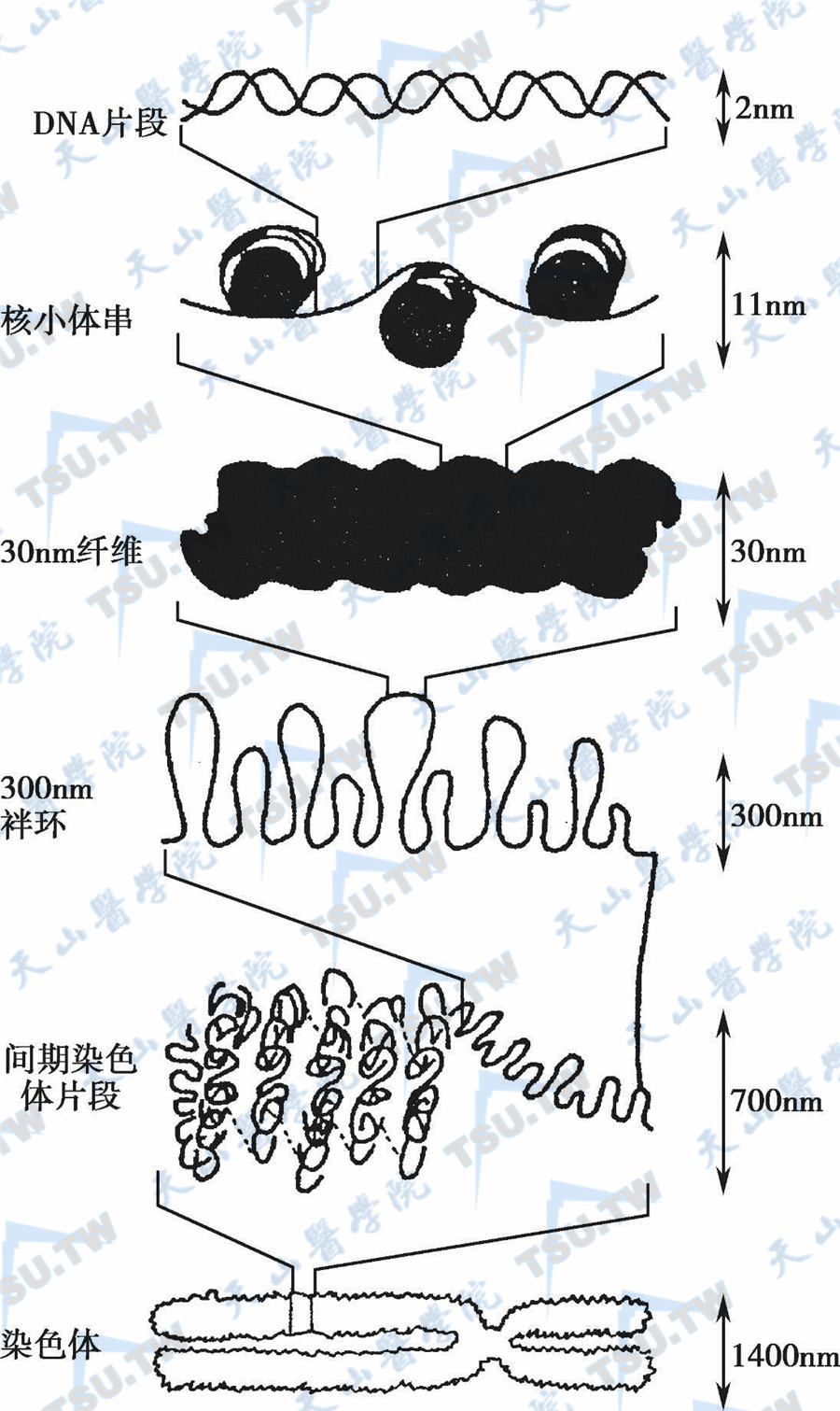
核小体由核心颗粒和连接区组成,核心颗粒的核心各由两分子H2A、H2B、H3和H4等4种组蛋白构成的八聚体及其围绕周围的DNA组成,直径11nm,这段DNA称为核心DNA,约含140个碱基对,围绕核心外周1.75圈。两个核心间的DNA链称为连接区,长约60bp。组蛋白H1位于连接区的DNA表面。连接区的DNA长度差异较大,短者8bp,长者114bp。核小体通过DNA分子串联,形成染色体的一级结构。在细胞核内,通过核小体结构,DNA长度被压缩至原来的1/37。核小体链进一步螺旋化,每6个核小体组成1个螺旋,形成外径为30nm的螺线管;DNA长度又被压缩了1/36。螺线管是染色体的二级结构,进一步螺旋化而形成直径为200nm的圆筒状结构(超螺线管),此时DNA长度又被压缩了1/40。超螺线管是染色体的三级结构,由超螺线管再缠绕折叠形成有丝分裂中期的染色体,DNA被压缩了1/5,此即染色体的四级结构。中期染色体由两条染色单体构成,直径约1400nm。这样,经过多级包装,DNA的长度被压缩至原来的近万分之一。
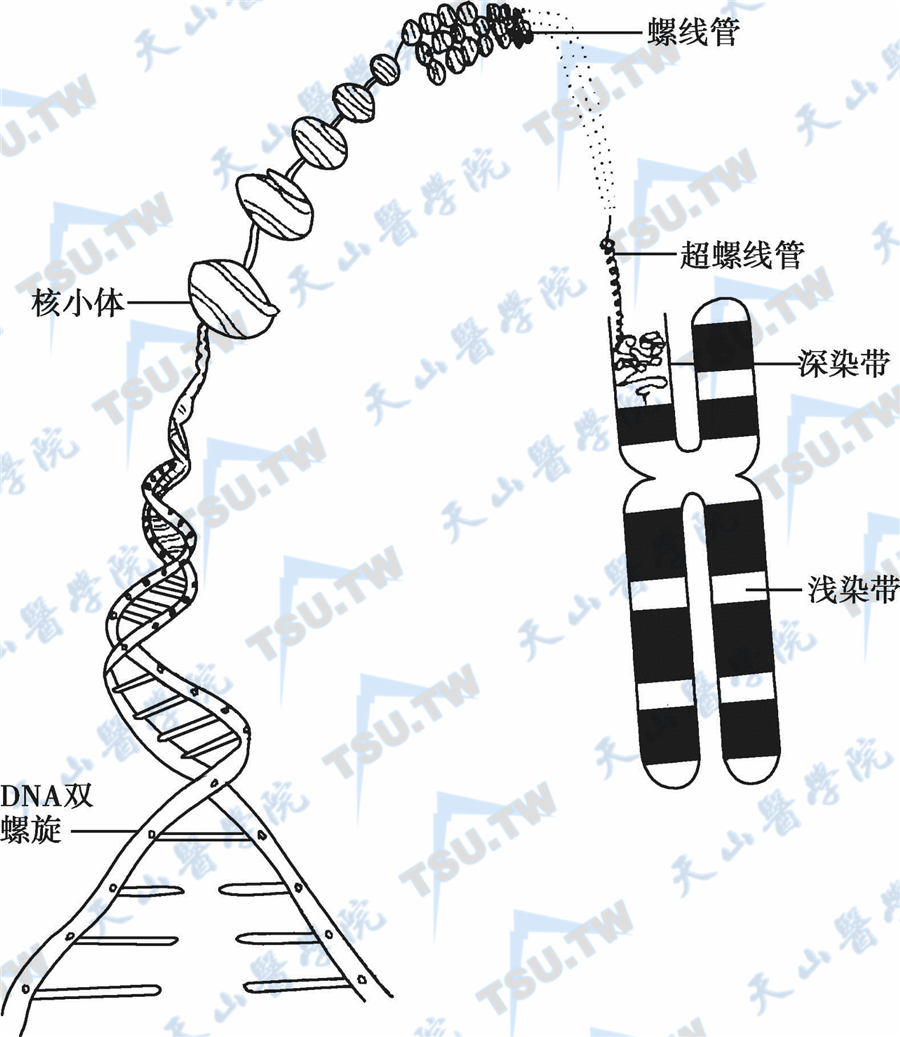
染色体结构(从核小体到染色体)
有关染色体结构的“袢环模型”认为,核小体进一步包装成直径30nm的染色体纤维,再折叠成袢环结构域(domain),袢环再经数次折叠盘曲而最终形成染色单体(下图)。染色体的这种结构有利于纺锤丝将染色体平均分配到两个子代细胞中,并保证不在此过程中断裂,有利于准确、高效地进行DNA复制和基因转录。
染色体的袢环模型
人类染色体数目46条。其中22对(44条)是常染色体(autosome),另2条与性别有关,称为性染色体(sex chromosome),女性为XX,男性为XY。
细胞分裂间期核染色体可见浅染带
异染色质是高度卷曲紧缩的染色体,一般属于不转录的DNA部分;而常染色质是伸展松散的DNA部分,正在进行活跃的转录。在细胞分裂期,染色质卷曲、螺旋化和凝缩,并逐渐表现出分裂期染色体的形态特征。在细胞分裂中期,染色体的凝缩程度最高,形态最清楚,故常采用此期的细胞染色体进行观察。中期染色体由着丝粒(centromere)相连接的两条染色单体构成,将染色体分为长臂(q)和短臂(p)。着丝粒附着在纺锤丝上,细胞分裂时牵引染色体向细胞两极移动。
利用各种技术展示不同染色体的特征,染色体可被有效识别。1960年,由第1届国际细胞遗传学会议制定的Denver体制是识别和分析人类染色体的基础。根据Denver体制,46条染色体按其长度及着丝粒位置分为23对,按顺次分为1~22号,并分为A、B、C、D、E、F、G等7组;其中男女性别所共有的第1~22对是常染色体,与性别有关的X染色体归入C组,Y染色体归入G组。非显带染色体技术能将染色体分组,但不能区别各组内染色体;而染色体显带技术利用各种方法处理染色体,使其沿纵轴方向显示宽窄不同的带型。显带技术进一步改进,产生了高分辨显带染色技术,它应用细胞同步化制片技术及改良的染色技术,使染色体显示出更丰富的带型,有利于准确识别每条染色体的区带,发现染色体异常。
