
一、受体变构效应使钙通道开放并激活蛋白激酶与信号分子磷酸化
钙调节蛋白(calmodulin)可改变蛋白的空间结构,增强酶的催化作用。有些激素以受体为介导,兴奋G蛋白,激活细胞膜磷脂酶C,继而使磷脂酰肌醇裂解为三磷酸肌醇(inositol triphosphate,IP3)和二酯酰甘油(diacylglycerol,DAG),后二者均为第二信使,可将激素等细胞外信息传递到细胞内。DAG可激活蛋白激酶C,使蛋白磷酸化,IP3可使细胞内质网和线粒体释放Ca2+。蛋白激酶C与Ca2+耦联扩增了激素作用(下表)。
表1:膜受体的特性比较

二、信号蛋白磷酸化引起系列生物反应
肽类激素信号在受体后(细胞内)的转导通路很多,归纳起来主要有:
- 第二信使介导的信号通路,其中又包括AC 酶-cAMP-蛋白激酶A通路、磷脂酰肌醇-Ca2+信号通路、二酯酰甘油(DAG)-蛋白激酶C通路、cGMP-蛋白激酶C通路、一氧化氮合成酶-一氧化氮通路等;
- 含酪氨酸激酶受体-信息转导蛋白-有丝分裂原活化蛋白激酶信号通路;
- 细胞因子-信息转导与转录活化(signal transducer and activator of transcription,STAT)蛋白信号通路;
- 第二信使-蛋白激酶-DNA基因表达信号通路等。对具体的激素来说,可能仅通过某一通路或通过1个以上的信号通路转导信号和级联扩增信号。
三 、膜受体存在信号对话现象
肽类激素、生长因子和细胞因子的膜受体结合具有很高的特异性(少数激素的分子结构类似或受体结合区的构象相同或相似,存在一定程度的交叉结合现象),但受体后的信息转导途径往往是共有的或交叉的,数百种激素的受体后信息转导通路至多也不过数种或10多种,所以受体后的信息传导系统相对受体结合特异性来说是低的,而且GPCR、RK、RLK和RLGIC等各受体后转导系统[如ras/raf/ MAPK/ERK激酶(MEK/MAP)级联反应系统]之间又相互影响或相互联系,这些都是发生受体后信息对话(cross-talk)的原因。
四、 受体后级联反应扩增信号
如上所述,作为激素信号传递的效应体(第二信使)很多(如cAMP、cGMP、Ca2+、IP3、DAG等)。它们是由酶(腺苷或鸟苷酸环化酶、磷脂酶A2或磷脂酶C)或离子通道等来反映的,而且还必须通过中介物-信号转导物(如G-蛋白)来传递。激素信号由细胞外向细胞内的传递特点是特异性高和信号级联扩增(cascade amplification,下图)。此外,各信号传递通路之间常相互影响。根据受体和效应体的差别,可将激素受体的效应体系统分为6类,见表2。激素对受体活性的调节为降调节。配体减少,受体有部分升调节,但代偿不全。例如,胰岛素抵抗(高胰岛素血症)时,受体的降调节加强,受体后通路减弱,使胰岛素的生物活性进一步下降。激素特异性信号传递产生级联扩增效益,作为激素信号传递的效应体很多(如cAMP、cGMP、Ca2+、IP3、DAG等),而且必须通过中介物-信号转导物(如G蛋白)来传递。
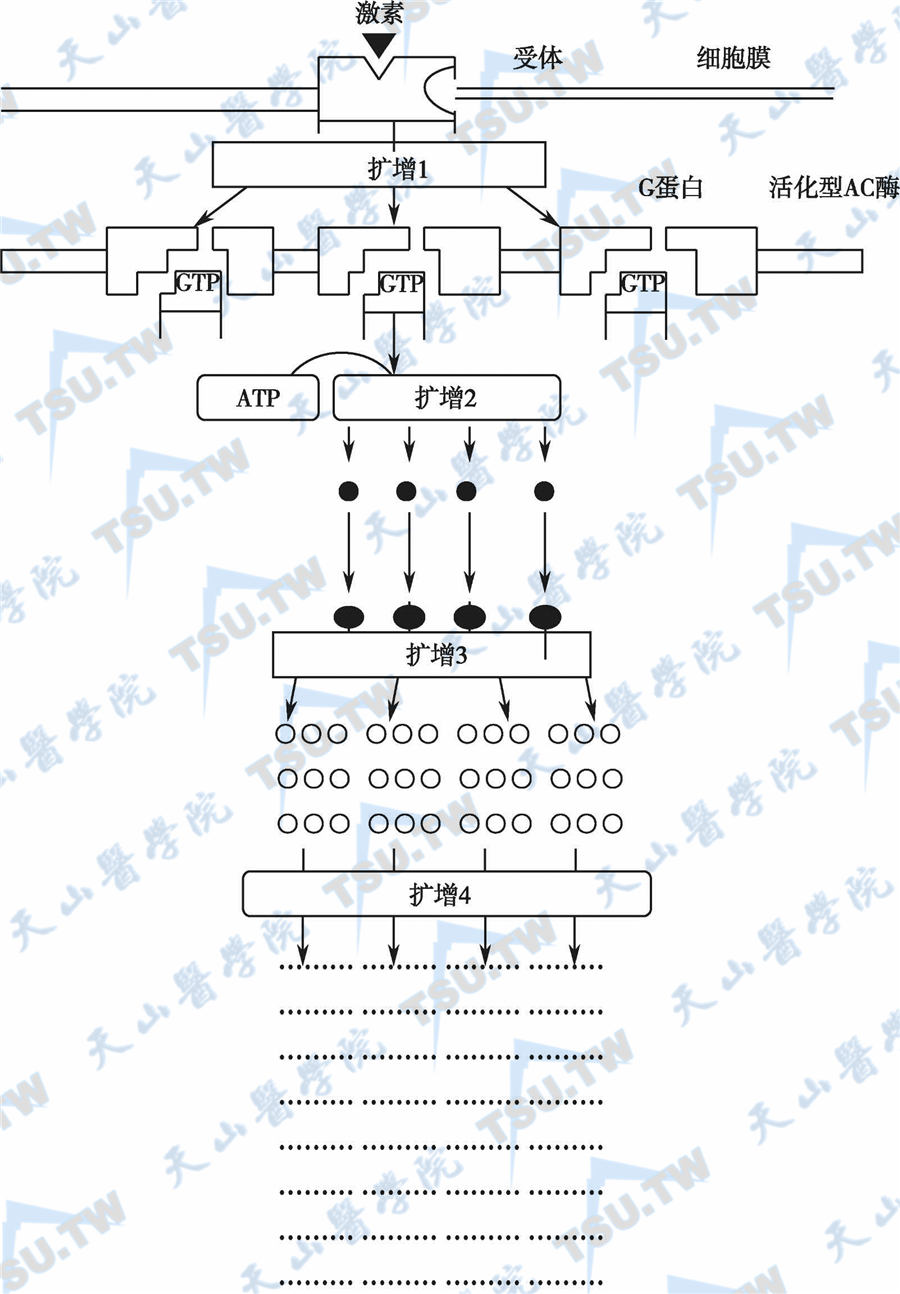
G蛋白偶联的激素受体信号级联扩增反应
注:扩增1:一分子激素活化-分子膜受体,后者可激活数个G蛋白;扩增2:一分子AC酶激活后可产生多个cAMP分子;扩增3:一分子cAMP激活一分子蛋白激酶,蛋白激酶磷酸化可激活多个酶X;扩增4:一分子活化型酶X可产生酶X的许多产物●:cAMP;●:蛋白激酶;○:酶X :酶X产物
:酶X产物
表2:膜受体/效应体类型

注:ACTH:促肾上腺皮质素;LH:黄体生成素;HCG:绒毛膜促性腺激素;FSH:卵泡生成素;GLP-1:胰高血糖素样肽-1;PTH:甲状旁腺素;IGF-1:胰岛素样生长因子-1;EGF:上皮生长因子;CSF-1:集落刺激因子-1;FGF:成纤维细胞生长因子;GH:生长激素:LHRH:黄体生成素释放激素;TRH:促肾上腺皮质激素释放激素。
必须指出的是,在激素与受体结合前,激素的结合蛋白对调节其浓度起了重要作用,它可进一步稳定并随时调节激素与受体结合的量。例如,GH/GHR/GHBP/IGF/IGFBP/ IGFBP酶组成反馈调节环,而IGFBP-3又被前列腺特异性抗原(prostate-specific antigen,PSA,即IGFBP酶)裂解,由于IGFBP-3与IGF的结合亲和性高于IGF受体,因此IGFBP-3调节IGF(GH)的可用性和生物活性的能力很强。一般来说,激素受体与受体后信号之间无反馈调节,其生理意义是确保激素作用的正常表达。当受体作用减弱后,受体后的信号扩增作用减弱,生物活性下降。
受体经糖化(saccharification)或酰化(acylation)而形成二硫键或裂解为成熟受体蛋白,受体合成或降解速度可引起膜受体数目的变化。多数情况下,激素通过受体数目减少(降调节,down regulation)来调节激素活性。基础状态下,受体在膜上呈弥散性分布。当受体与激素结合后,弥漫分布的受体向受体-激素复合物处聚集,并形成二聚体,继而可形成多聚体。受体-激素复合物由胞膜的包裹口(coated pits)内陷并移至胞质内。在胞质内形成内饮体(endosome;受体小泡,receptosome)。内饮体与溶酶体融合后被消化、分解,或经过出胞作用(exocytosis)进入受体的再循环途径,而激素被降解灭活。
