
在胚胎和出生后的乳腺发育过程中,生长因子介导的上皮-间质相互作用发挥了关键性作用。胚胎乳腺的发育和其他皮肤附属物如牙胚、毛发、毛囊等的发育过程是类似的,而乳腺始基的发育是上皮与间叶持续相互作用的结果。
乳线的形成
在胚胎发育的第10~11天,乳腺始基表现为在躯干的两侧上皮增厚,称为乳线(milk streak, milk line)(图1-13A)。乳线分子定义为Wnt-10b mRNA的表达。
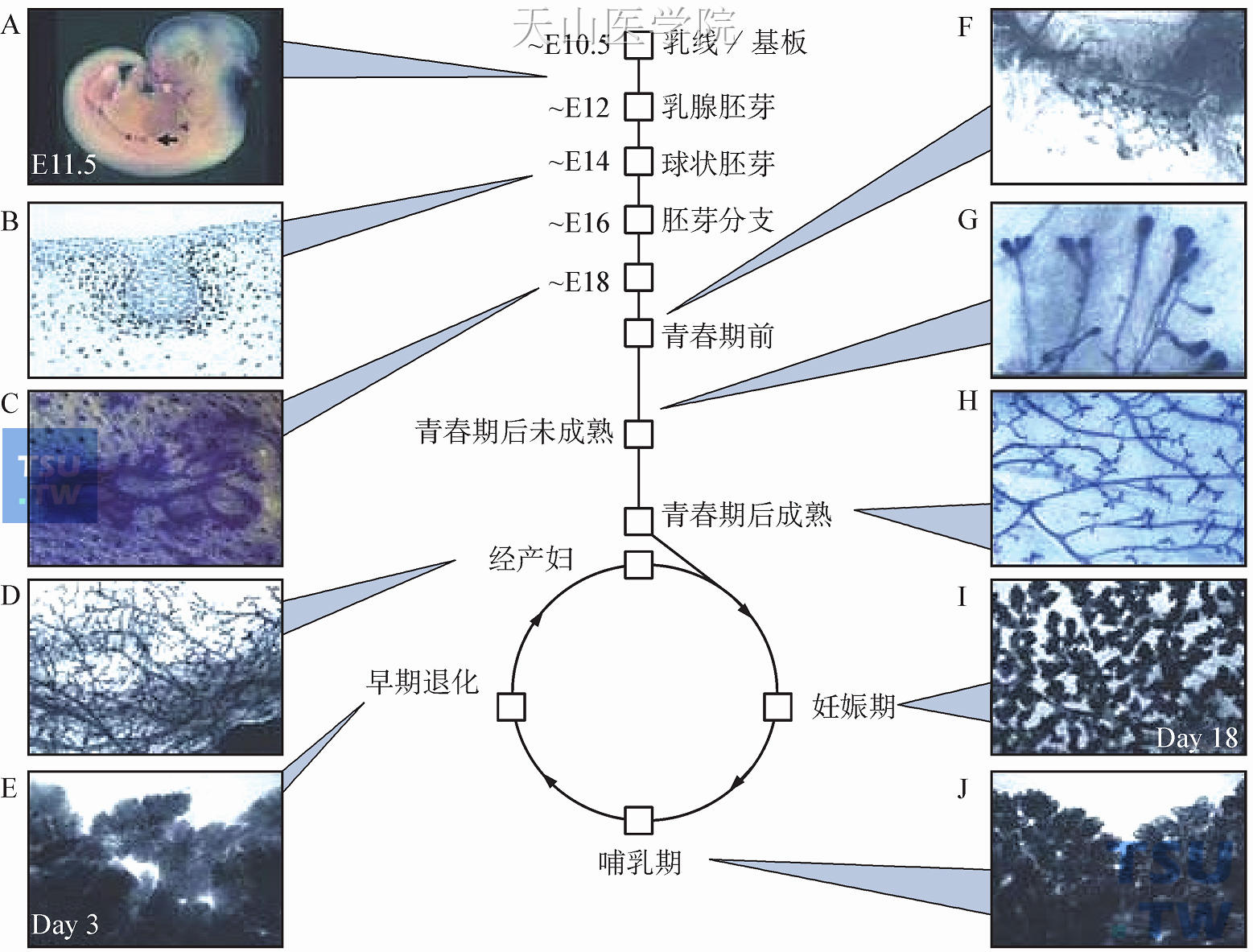
图1-13 乳腺发育综览
注:乳腺发育可分为线性期(胚胎发育的原始导管成熟)和与妊娠、哺乳、退化相关的循环期。A可确定胎儿Wnt-10b表达(原位杂交)的胚胎第11.5天的乳线和乳腺始基(箭头所示),可见基板3、4。B.胚胎第14天的球状期,球周致密的乳腺间质和雄激素受体(见染色处)优先表达。C.胚胎第18天的胎儿乳腺始基表现出轻微的分支。D.完全退化的经产妇乳腺,在形态上跟成熟未婚女性的乳腺相似。E.乳腺退化的早期(3天),此时发生明显的细胞死亡,早期退化乳腺和哺乳期乳腺在形态上仅有轻微差异。F.青春期前的乳腺表现出从出生到青春期的典型形态。G.青春期后,未成熟乳腺表现为明显的终端细芽和简单的分支模型。H.青春期后,成熟乳腺可见完全分叉的导管和导管终末变钝的末端。I.妊娠期(腺体来自妊娠18天的大鼠),腺泡很明显,但没有扩张。J.哺乳期,腺泡很大并扩张,几乎完全充满了脂肪垫。
基板的诱导
在大鼠,有5对外胚层基板或原基(在人类是1对)同时开始形成,并在1天之内这些基板形成鳞状上皮细胞,后者在形态学上与周围上皮截然不同。致密乳腺间叶细胞是由2~3层排列紧密的成纤维细胞组成,并包裹正在发育的乳腺胚芽,后者在胚胎发育的第13天出现。间叶细胞的功能是维持上皮胚芽的生长和调控睾丸激素刺激下雌雄异型的发生。晚期较致密乳腺上皮细胞存在于脂肪垫中,由前脂肪细胞构成。到胚胎发育的第14天,乳腺间叶细胞表达相对高水平的雄激素受体(图1-13B)。在雄性小鼠,睾丸分泌的雄激素导致乳腺上皮胚芽活性减退。在雌性小鼠,原基继续缓慢生长到胚胎第16天,这时细胞增生加速,乳腺上皮的乳芽开始长入周围的脂肪垫,开口于乳头(图1-13C)。出生时,连接乳头的主导管形成,包含12~15个含导管上皮的小分支(图1-13C),其存在于始基中直到出生(图1-13F)。
尽管调节胚胎乳腺发育的确切分子机制还有待于阐明,但已经发现这个过程涉及越来越多的转录因子和局部生长因子。某些基因为乳腺发育所必需,转录因子p63便是最早发现的基因之一,它是p53基因家族的成员。P63蛋白产物对于哺乳动物表皮的发育极为重要。p63基因缺失小鼠所有的复层扁平(鳞状)上皮细胞及其衍生物均缺如,包括乳腺组织。p63由两种不同的启动子调控转录,从而导致6种不同的蛋白异构体出现,它们具有活性功能或显性失活。特定的p63异构体在诸多方面有着不同的作用,不但表现在维持上皮干细胞数目方面,而且对于细胞分化和肿瘤也有一定意义。用一种能识别所有这些异构体的抗体,可发现早在胚胎第16.5天的乳腺胚芽中p63即有表达。同时,在邻近上皮细胞的基底层也有表达。在胚胎发育的这一阶段,乳腺胚芽中也发现有角蛋白14(CK14)的表达。Np63作为p63表达形式之一,可以调节细胞核内β-连环蛋白(β-catenin)的表达,从而影响经典的Wnt信号通路。Tap63α作为一种异构体,已经被报道能增加成纤维细胞生长因子受体-2(FGFR-2)的表达,这对乳腺基板的构成也是非常重要的。
胚胎间叶细胞与上皮之间的相互作用对乳腺胚胎发育至关重要,这种相互作用由FGF家族(如FGF-10)及其受体酪氨酸激酶(如FGFR-2b)介导。FGFR-2-IIIb是FGFR-2的一个特殊异构体,在胚胎第11~12天的乳腺基板中表达。若小鼠上皮细胞中的FGFR-2-IIIb受体表达缺失,或间叶细胞周围的配体FGF-10表达缺失,均可导致基板4、5诱导缺失。集聚在乳腺萌芽周围的间叶细胞还表达FGF-7,即FGFR-2-IIIb的另一种配体。但是缺失FGF-7的小鼠不会出现乳房缺陷。
在应用报告基因的转基因小鼠(TOPgal)模型中,通过β-连环蛋白/T细胞因子(TCF)调控的β-牛乳糖报告基因的表达,β-连环蛋白信号通路得以呈现,因此最早在胚胎第10~11天即可检测到β-牛乳糖阳性细胞。此模型显示Wnt信号途径在乳腺胚芽形成过程中的重要性。Wnt-10b及其下游靶点Lef-1(即Lef/TCF转录因子家族成员之一)均是乳腺基板的早期标记。Lef-1基因的缺失导致乳房障碍形成。某些器官的发育依赖于其诱导的间叶-上皮相互作用,而这些器官的发育也会受到影响,如牙齿、胡须、头发等,这与p63无效基因的胚胎类似。Wnt信号途径功能的重要性还可以通过一种Wnt信号抑制剂Dickkopf-1的表达来证实,它来自转基因鼠CK14的启动子,能导致乳腺胚芽的缺失。因此,Wnt通过β-连环蛋白的信号转导对于乳腺基板的形成十分关键。
研究表明,胚胎发育过程中若缺失Gli3的转录抑制功能,将导致乳腺基板3和5的形成障碍。对于基板3,Veltmaat等的研究表明,Gli3功能对早在胚胎第10.5天胎儿胸节的脊柱轴下出芽是必需的,其可以诱导FGF-10的表达。正如以上所述,表外胚层通过FGF-2b受体接收FGF-10信号,并诱导乳线上Wnt-10b表达。缺失Gli3将导致FGF-10表达减少,以及与乳腺基板3对应的区域Wnt-10b诱导失败。因此,对于乳腺基板3,Gli3功能对于体节乳腺上皮细胞的形成是必需的。但是,在上皮细胞自身早期基板生长中Gli3似乎并不是必需的。
乳腺新芽的延伸和乳腺脂肪垫初期的扩展
上皮细胞与间叶细胞相互作用的信号途径涉及甲状旁腺素相关肽(PTHrP),它在胚胎发育第11.5~18天的萌芽上皮中表达,并且通过G-蛋白耦联受体PTHrPR1作用于周围的间叶细胞,诱导密集的乳腺间叶细胞形成。小鼠若缺失任何一种配体或受体,乳房就会在接近胚胎第15天时停止发育,分支延伸失败。缺乏PTHrP信号导致乳腺上皮细胞回复至普通上皮细胞。乳腺上皮细胞和间叶细胞表达的配体与受体的相互作用可以调节信号途径,而这些信号途径对胚胎乳腺发育十分重要。
对后天乳房发育十分关键的还有雌激素受体(ER)和孕激素受体(PR),两者在胚胎乳腺均有表达。ER的两种异构体在胚胎第12.5天小鼠晶胚的乳腺间叶细胞中可检测到,而PR则在乳腺胚芽的上皮细胞中表达。然而,目前还没有关于敲除ER或PR异构体会对小鼠胚胎乳腺表型产生何种影响的报道。
人类胚胎乳腺发育缺陷
尺骨-乳腺综合征:T-box基因为TBX3基因的自发性突变,可导致人类尺骨-乳腺综合征。这是一种显性发育异常,表现为前臂和顶浆分泌腺体的发育异常。TBX3同源体缺失的小鼠突变型,也显示乳腺诱导的缺失,以及上肢和其他畸形。TBX3突变小鼠还缺乏Wnt-10b和Lef-1的表达,提示这一转录因子可能位于Wnt信号途径的上游。
多乳头和多乳房:多乳头和多乳房是人类常见的出生缺陷,发生率5%。多数病例属于偶发,但也有与遗传性相关的例子。奇怪的是,许多遗传形式伴有其他的发育异常,尤其是上肢和手指(足趾)缺陷(并指或多指)、颅骨缺陷(唇裂、腭裂)及肾异常。一些遗传形式跟某些癌症(肾腺癌、Wilms瘤)的发生率增加有关。但偶发性或遗传性多乳头或多乳房的特异性变异还不清楚。Simpson-Golabi-Behmel综合征由X-连锁基因引起,伴有磷脂酰肌醇蛋白聚糖-3(glypican-3,GPC-3)缺失,后者可以与胰岛素样生长因子-2(IGF-2)相互作用。
人们已经确定了许多多乳头和多乳房的大鼠模型。神经调节蛋白-3(neuregulin-3,Nrg-3)基因编码一种表皮生长因子(EGF)超家族的分泌型配体,其点突变不仅导致乳腺基板3高频缺失,还引起多乳头。Nrg-3在间质细胞表达,并通过与erbB-4受体结合从而将信号转到表外胚层。某些其他的Nrg和erbB受体以发育调控和空间限制模式也在胚胎乳腺和相关间质表达,这表明上皮和间质涉及信号网络的一种复杂的相互作用。
