
受精过程是精卵相互作用最终形成受精卵的复杂过程,包括对透明带的黏附、识别和穿透、精-卵识别、精子的穿入和精卵的融合等。研究表明,精子的受精能力是在附睾运行和贮存过程中获得的,是附睾精子成熟的核心。附睾精子受精能力获得的研究是Bedford和Orgebin-crist于1967年首先在兔子上进行的。他们采用结扎方法使附睾精子不能循附睾头体尾进行运行,而只能停留在附睾近中段。虽然近中段附睾精子能存活,但却无受精能力。随后上述两位学者和其他科学工作者对绢猴、猪、羊、小鼠、大鼠、田鼠等附睾精子进行了系统研究;在人也发现大部分附睾精子进入到体尾部的过程中获得了受精能力。而在附睾管阻塞或缺失时,近中段部分的精子仅一小部分具有受精能力。至此,对包括人在内的附睾精子受精能力获得的部位基本清楚。
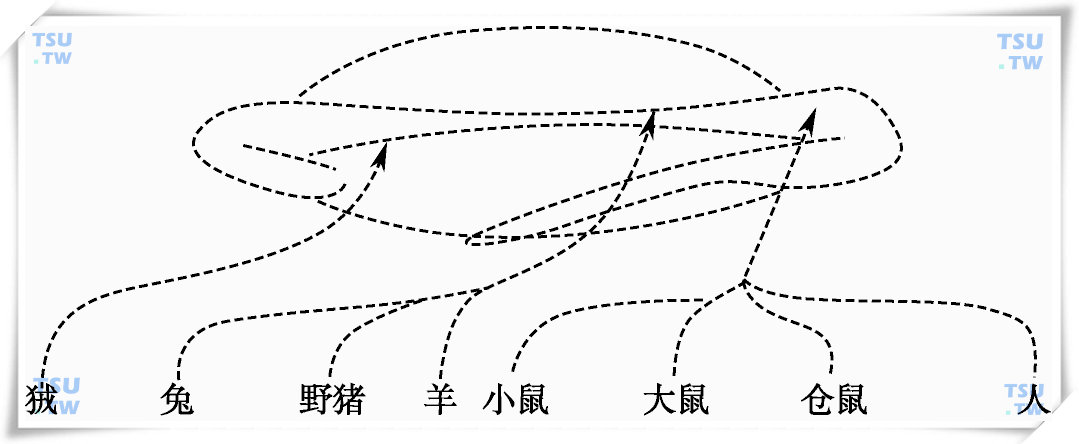
人及动物附睾中精子受精能力获得部位图示
为了使卵子受精,精子必须获得运动和受精能力,从而在女性生殖道内运行,在女性生殖道精子进行获能、识别、黏附和穿透透明带、精卵的识别和融合,从而形成受精卵。附睾精子受精能力的获得已得到证实,但受精这是一个十分复杂的过程,涉及很多环节。因此附睾精子受精能力的获得也决不会是单一的变化。近年已有不少研究工作,特别是一些分子生物学的研究工作正在揭示出附睾精子受精能力获得和发育的机制及其本质。
精子受精能力的获得和发育首先表现在精子对卵丘细胞层的穿越作用。长期以来,人们却忽略了精子对卵丘细胞层的作用。近几年来研究发现,精子膜表面的PH-20蛋白具有透明质酸酶的活性,可使卵丘细胞很快散开。如果PH-20被透明质酸酶抑制剂和抗氨基端活性部位的抗体封闭,精子就无法穿过卵丘细胞层。虽然PH-20蛋白是在精子发生过程中形成,并分布于精子头部,但在附睾精子成熟过程中PH-20蛋白的定位发生了变化,主要定位于精子头后部质膜和顶体内膜,后一部位PH-20蛋白的量是前者的两倍,浓度明显增高。PH-20蛋白在精子膜上的重新分布和浓度变化在受精起始阶段有着十分重要的作用。穿越卵丘细胞后所发生的精子对透明带的黏附和识别。这种黏附和识别主要是精子表面的糖基和透明带的糖基结合蛋白IP3之间的结合。研究表明,精子对透明带的黏附和识别能力是在附睾中获得的。有人将小鼠和羊附睾头部精子与同种卵细胞孵育,发现均不能固定于透明带,而附睾尾部精子却能较好地固定于透明带。附睾头部精子不能固着于透明带的原因是由于精子未成熟,缺乏对透明带的黏附和识别,这可能与附睾头部上皮细胞分泌的酸性糖蛋白有关,将该蛋白与未成熟精子一起孵育能增加精子固着于卵子透明带的能力。
精子黏附并穿越透明带后,精子需要和卵细胞质膜融合才能完成受精。精子和卵细胞表面的蛋白质复合物通过结合和融合进而完成质膜的融合。一些附睾分泌的精子结合蛋白参与了这个过程。
精子膜融合蛋白Fertilin在精子膜蛋白与卵细胞膜的融合中可能起着关键作用。Fertilin主要位于精子头后部质膜或赤道板区,它由α和β亚单位构成。α亚单位是在睾丸精子发生过程中被酶切,而β亚单位则是在附睾运行过程中成熟(下图)。
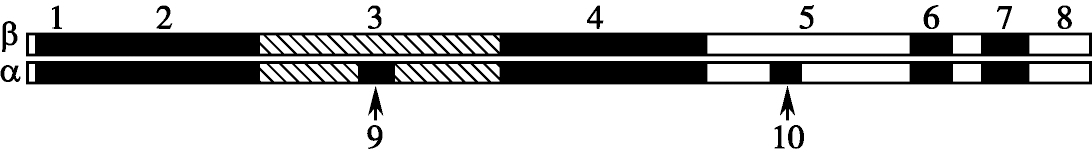
Fertilin蛋白α和β亚基结构图
虽然α和β亚单位的结构和氨基酸序列相近,但是它们的功能则有明显的差别。荧光标记的重组β亚基正好与卵母细胞表面的精子结合位点发生作用,体现为整联蛋白配体区顶端的TDE三肽结构与卵母细胞质表面的型整联蛋白结合,一旦去除整联蛋白的配体区,则β重组亚基的结合能力丢失。另外发现敲除编码β亚基整联蛋白第14外显子区,子代精子在体外黏附和融合去透明带卵母细胞的能力明显下降。
酸性附睾糖蛋白(DE/AEG)在精子膜与卵细胞膜的融合中也起着重要作用,它包括附睾蛋白DE、酸性附睾糖蛋白AEG、唾液酸蛋白和相对分子质量为32 000蛋白等。其中附睾蛋白DE最早在大鼠发现,为相对分子质量37 000糖蛋白,由附睾近段合成分泌并结合到精子表面。DE蛋白最初位于精子的顶体区。在获能和顶体反应后迁移到了赤道区,间接免疫荧光证实,在小鼠卵子质膜表面存在DE蛋白结合位点。重组人ARP蛋白的抗体可以阻止人精子穿越去透明带仓鼠卵,但不影响精子的运动能力、顶体反应能力以及与卵透明带的结合能力。同时,在人的卵细胞膜上也发现有ARP的结合位点,提示ARP参与了精卵融合过程。此外,人附睾蛋白SOB2、FLB1、gp20、Ecad等也参与了精卵融合过程。
长期以来,人们十分关注附睾上皮合成的蛋白质与附睾精子表面结构的关系,关注在受精过程中的重要作用。不少学者将附睾上皮合成的并参与精子表面结构的蛋白质称为成熟抗原,并认为与精子受精能力的获得有关,在受精过程中发挥作用。
Kirchhoff等用分子生物学方法,研究了人附睾蛋白及其精子成熟过程中的作用。他们主要研究了六种附睾分泌蛋白,其中有些蛋白质可能与精子成熟及精子受精能力获得相关。
一、HE1可能与胆固醇转移有关
HE1编码的人附睾蛋白为分泌性糖蛋白,并在附睾尾液中积累,也大量出现于精液中。这种蛋白质的功能主要涉及胆固醇的转移,被认为是一种“去能因子”,旨在使精子在附睾运行和贮存过程中维持精子膜胆固醇的量,而在进入女性生殖道获能过程中,胆固醇从精子膜上漏出,因而精子膜胆固醇被稀释和散失。
二、HE2是起源于附睾头部的精子表面抗原
HE2cDNA编码人附睾近侧一种小分子分泌性糖基多肽,它的mRNA具有高度的附睾特异性,尚未在其他人类组织中发现。推测HE2是一种新颖的人精子表面抗原,可能与配子融合有关。
三、HE4的cDNA编码
一种来自于人远端附睾上皮的小分子量的酸性蛋白HE4抗多肽血清与射出精子的整个表面反应。HE4的二级结构与精子膜上的麦芽凝集素的结构很相似,后者是一种植物凝集素。精子获能的一个重要过程是去除精子膜上的这层保护性蛋白衣。而精子膜上的这层表面蛋白衣是精子的附睾运行和成熟过程中或射精时沉积上去的。因此,HE4可能是精子的一种新颖去能因子,并认为可能与获能和受精过程有关。
四、HE5是分离筛选出的重要的人附睾基因产物
HE5cDNA和淋巴细胞的CD52的cDNA相一致,编码出一种小分子糖磷脂酰肌醇GPI。原位转录杂交显示出很大部分的附睾mRNA来自于附睾管腔上皮,CD52抗体与部分男性生殖管道上皮发生反应,特别是与附睾尾部上皮和附睾尾液发生反应,和附睾精子而不是和睾丸精子发生反应。这个结果与以下观点一致,即抗原仅由睾丸后生殖管道上皮细胞产生,并与它的GPI-锚着物一起完整地分布于管腔面,并且通过它的GPI-锚着物结合到精子膜上。已知大鼠CD52是其主要成熟抗原或精子膜糖蛋白SMemG,表明它是一种完整的精子膜蛋白,发生于远端附睾精子表面,而这与附睾精子获得成熟的生理特征相吻合。至今人们已知道人淋巴细胞CD52-连接的碳水化合物结构,包括了大量岩藻糖,甘露糖核心相连的唾液酸多聚乳糖单位。在附睾远端,精子膜“糖衣”也是一种碳水化合物结构。同时发现这类碳水化合物的许多单克隆抗体能和精子结合,并能抑制受精。在一些不孕妇女中也能分离出这种类似的抗体。应当指出,附睾精子受精能力的获得和发育机制的研究还很不充分,有待深入。
