
顶体及顶体酶
哺乳动物精子顶体是一个膜结合的帽状结构,覆盖于精子核的前面。顶体由位于前面的顶体帽和位于后面的赤道板两部分组成。
精子获能后,精子膜不稳定性增大,去除了抑制顶体反应的因子,精子就开始顶体反应。哺乳动物卵子透明带具有诱导精子发生顶体反应的能力。在小鼠,ZP中的肽链具有这种功能。海胆卵子胶质外衣上的多糖可触发海胆精子顶体反应。Dan提出,当海胆卵子胶质外衣上的多糖与精子膜上的蛋白质受体结合,便引起精子质膜的构象改变,使膜上的通道短暂开放,大量Ca2+内流。胶质外衣上的多糖还激活精子膜上的质子泵,将细胞内H+泵出并使Na2+内流,细胞内的pH值升高与Ca2+浓度增加一同引发了顶体反应。最初精子顶体外膜与其相贴的精子细胞膜发生众多的点状融合,形成许多膜包小泡,由于顶体外膜的破损,顶体内容物外溢,直至将顶体内膜完全暴露。顶体反应完成的标志是顶体外膜与精子细胞膜的完全融合(下图)。
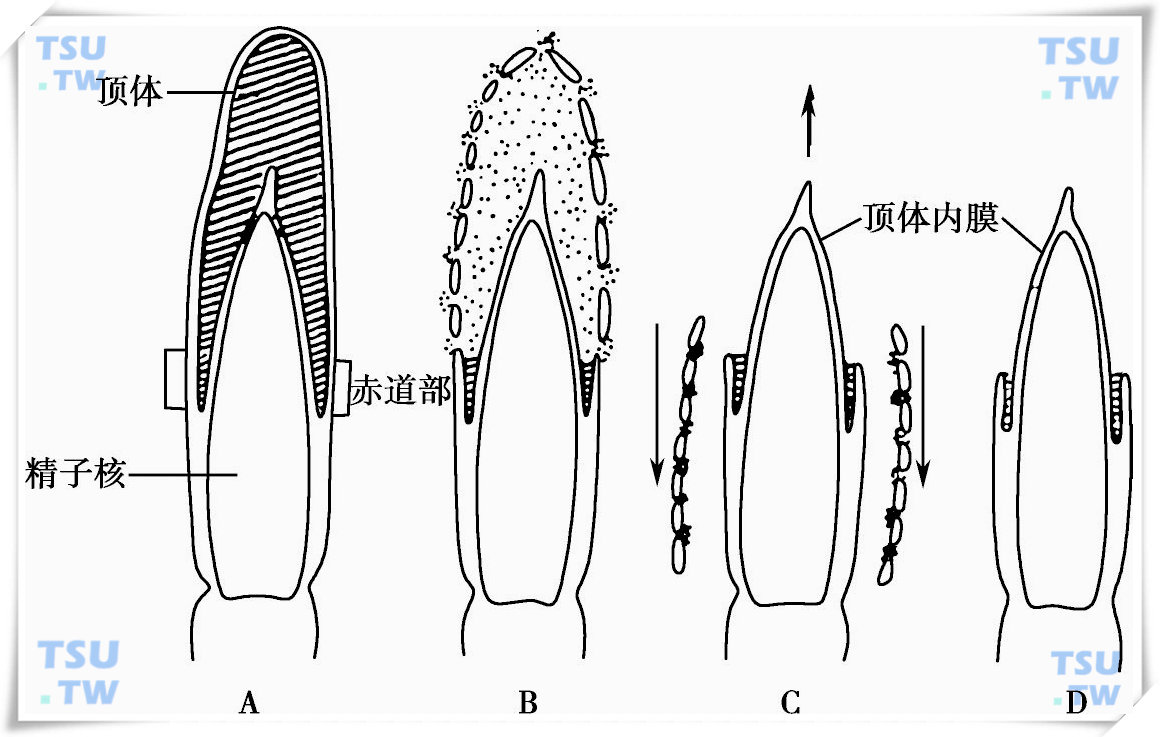
顶体反应过程图解
Ca2+是激发哺乳类精子顶体反应的必需因子,顶体反应适宜的条件有:温度23~40℃,pH7.2~7.8,渗透压230~343mOsmol/kg。加入血清白蛋白可加速Ca2+内流而促进顶体反应。在生理条件下,附睾液中Ca2+浓度低而输卵管中浓度高,显然有利于顶体反应。顶体含有相对分子质量为59 000的顶体酶复合系统,其中包括许多酸性水解酶,故将顶体视为一个特化的溶酶体。已发现顶体所含的酶至少有20种以上,其中与受精有关的主要酶类有:
- 透明质酸酶(hyaluronidase),位于顶体的边缘与前部。人精液中抽取的透明质酸酶可使兔卵丘细胞分散并使透明质酸解聚,这显然有利于精子靠近并与卵子结合。人工合成的此酶抑制剂58D/K加入精液可抑制受精作用。
- 放射冠分散酶(corona removing enzyme),是一个含脂的糖蛋白,此酶可移除卵子表面的放射冠。
- 顶体蛋白酶(acrosin),亦称顶体素,主要位于顶体内面及赤道部位,作用类似胰蛋白酶,故又称类胰蛋白酶。顶体蛋白酶具有氨基肽酶、酯酶及蛋白酶多种活性,并对精氨酸键有高度选择作用。现在认为,顶体蛋白酶起初与顶体内膜紧密结合,此时无活性,是一种顶体素酶原(proacrosin)。当Ca2+大量内流时,酶原激活成为可溶性顶体蛋白酶,进而可使透明带分解,这在受精中起十分关键的作用。天然的和人工合成的顶体酶抑制剂可抑制精子穿过透明带。
- 精子神经氨酸苷酶(SN)是与顶体结合最紧密的酶。一般的神经氨酸苷酶可以水解涎酸连接到次末端糖基的两类键(2→3,2→6),而SN只能水解2→6键。有关SN在受精中的作用还不大清楚,有人认为SN可以改变透明带结构,使其他精子不易穿入。下表列出了若干种顶体酶类。
已知的顶体酶类

总之,在输卵管内许多精子经历顶体反应释放大量水解酶,使卵丘细胞分散,放射冠移除。另外一些尚未发生顶体反应的精子得以与透明带接近,相互识别,继而也发生顶体反应,释放顶体蛋白酶,分解透明带,逐步穿入。用扫描电镜观察仓鼠体外受精过程,发现精子先在透明带上开一条很深的沟,然后精子头部斜插深入透明带。精子头部十分坚硬,加上获能精子拍击样摆动,使精子穿过透明带与卵细胞膜靠近。有依据提示,至少有一部分男性产生的精子是有缺陷的。这些精子不能完成获能并达到与引起精子内Ca2+升高的卵子相互作用的阶段,因而不能导致顶体反应。另外一些男性,无论是用卵子与其相互作用或者甚至使用钙离子载体A23187,其精子对Ca2+升高都不能产生反应。
荧光探针可用于观察人类精子的顶体反应。常用的荧光探针有三种,分别为:金霉素(CTC)、荧光素标记的甘露糖化的小牛血清(MAF)以及阿的平(QN)。研究结果表明,正常人射出的新鲜精液30~60min内,其未获能精子的顶体反应率低于10%,而且3种方法检测结果差别不明显。精子获能3h后,CTC和MAF法检测顶体反应率为12%,而QN法检测顶体反应率为18%(P<0.001)。加入钙离子载体A23187 1h后,CTC和MAF法检测顶体反应率为31%,而QN法检测顶体反应率为45%。从上述结果来看,CTC和MAF法相当吻合,并使我们能够从中看出精子获能以及Ca2+对顶体反应的影响。
顶体反应的生理学意义
一般认为,顶体反应所释放的透明质酸酶可消化卵子透明带外面的细胞外基质,并由精子携带的顶体酶促使精子穿过透明带。而且,只有经过顶体反应的精子才能通过透明带,并与卵膜融合。因此,Yanagimachi提出,顶体反应至少具有双重功能:①使精子得以穿过透明带;②使精子质膜与卵子质膜融合。但是,顶体反应的具体功能和意义还值得进一步探讨,例如,近期有研究表明,体外实验中,大部分小鼠精子在未接触到卵子透明带前即已经发生了顶体反应;而且,已经发生了顶体反应的小鼠精子,仍然可以使卵子受精。
顶体反应的相关蛋白
精子顶体反应的发生是通过精子膜蛋白与卵子透明带上糖蛋白间的相互识别完成的,有多种蛋白分子参与,如精子表面的胞外酶、细胞黏附分子和凝集素等。而卵细胞表面蛋白主要是组成透明带的糖蛋白ZP1、ZP2和ZP3,尤其是ZP3寡糖链起着识别信号的作用。
精子表面相关蛋白
β-1,4-半乳糖基转移酶Ⅰ(GalTⅠ):GalTⅠ能将半乳糖基转移至糖蛋白和糖脂上,但是分布在细胞膜上的GalTⅠ具有分子识别、黏附及介导信号传导的功能。GalTⅠ是最早报道的卵细胞结合蛋白。完整的膜GalTⅠ可以分为三个部分:胞外段、跨膜段和胞内段。胞外段作为凝集素,能够识别并结合末端为N-乙酰葡萄糖残基的寡糖链。胞内段具有信号传导能力,通过结构改变与G蛋白作用,传递信号。GalTⅠ与ZP3寡糖链识别并结合可使精子表面GalTⅠ聚集,激活耦联的G蛋白,引发顶体反应。然而,GalT缺失的精子仍然可以与卵子透明带结合,提示精卵结合过程中存在其他不依赖于GalT-ZP3的结合机制。
ZRK/hu9:在人和小鼠精子上都存在一种相对分子质量为95 000的酪氨酸激酶受体(RTK),位于精子头部顶体外侧质膜上,能够特异地与透明带糖蛋白ZP3结合,是公认的ZP3受体。小鼠的这类受体称为ZRK,人的则称为hu9。其大致可分为胞外段、单次跨膜段和胞内段三个部分。胞外段可以与ZP3的多肽链结合,胞内段含有酪氨酸磷酸化位点和Src蛋白结合位点。当精子与透明带结合后,其胞内段酪氨酸残基被酪氨酸激酶磷酸化,随之被含有SH2结构域的靶蛋白识别并结合,通过信号通路引起精子顶体反应。酪氨酸激酶抑制剂可抑制其Tyr残基磷酸化,进而抑制顶体反应。
甘露糖受体(mannose receptor,MR):MR属于C型凝集素超家族成员,是Ⅰ型跨膜受体,可以识别末端为甘露糖、岩藻糖、N-乙酰氨基葡萄糖的糖链。甘露糖受体定位于精子膜下层。当精子在女性生殖道内获能时,甘露糖受体随精子膜结构改变转向膜表面,与透明带上的游离甘露糖结合,快速诱导具有受精能力的精子发生顶体反应。
透明带黏合素(zonadhesin):透明带黏合素能与ZP3特异结合,是一个单次跨膜蛋白。羧基端位于胞内,较短。胞外部分相对较大,类似于嵌合蛋白,包含多个MAM结构、黏合素(mucin)结构、D结构和表皮生长因子(EGF)类似结构域。最近研究表明,透明带黏合素定位于顶体基质而不是质膜上,因此推测其在顶体反应后介导精子与透明带结合。
卵子表面相关蛋白
ZP3:ZP3是哺乳动物卵细胞透明带的重要组成成分,哺乳动物ZP3的同源性很高,相对分子质量约为94 000,具有O-连接和N-连接的寡糖链。ZP3的多肽结构共有424个氨基酸残基,分为信号肽序列、同源结构区、铰链区、精子结合区、成对的碱性氨基酸蛋白酶切位点、疏水区。目前研究表明,ZP3的寡糖链在精卵识别过程中起主要作用。包含4种精子识别必需的末端单糖残基:α-半乳糖、N-乙酰氨基葡萄糖、岩藻糖和甘露糖。在精子与卵透明带作用过程中,卵子表面成分作为精子表面受体的配体,起着信号分子的作用:①完成精卵识别。②诱发精子顶体反应。此外,ZP3对精卵结合后下游信号通路的调节也起着重要作用。
CD81:CD81在氨基酸序列上与CD9同源性很高。已证实CD9参与了精卵膜融合过程,而CD81也参与膜的融合。一般情况下,卵膜表面以及卵细胞周围的卵丘放射冠细胞都高表达CD81。实验发现,CD81双阴性的卵细胞(CD81-/-)不能诱导精子顶体反应。因此认为,在精子与卵子相遇前,卵丘细胞表面CD81与精子作用并参与了顶体反应的发生。
顶体反应的信号传导
顶体反应的发生是一个快速的过程,大多不是通过调节基因表达来完成,而是通过胞内已存在信号分子启动顶体与质膜的融合完成。目前认为,精子顶体反应中信号传导过程大致分为3个步骤:①精子膜表面受体聚集。这就要求精子完成获能,使质膜有良好的流动性;②通过受体介导激活信号通路;③在信号分子作用下,打开电压门控Ca2+通道,维持较高Ca2+水平。参与该过程的受体主要有G蛋白耦联受体、酪氨酸激酶相关受体。而从第二信使激活的角度看,主要有四条信号途径:G蛋白信号传导途径、酪氨酸激酶信号途径、丝/苏氨酸蛋白激酶途径和蛋白水解酶途径。
G蛋白耦联的信号传导
G蛋白是细胞内重要的信号分子,许多受体与其效应器之间都经过G蛋白介导。与受体耦联的G蛋白都是膜整合蛋白,由3个亚单位(α,β,γ亚单位)组成。α亚单位具有特异的GTP结合位点,有GTP酶活性,通过GTP-GDP的转换调节G蛋白活性。而G蛋白耦联受体在结构上有很大的相似性,都由一条7次跨膜的肽链构成,氨基端在胞外,羧基端在胞内,胞外部分以独特的空间结构共同构成配体结合结构域,胞内部分则可与G蛋白α亚单位结合。
活化的G蛋白α亚单位与β,γ亚单位分离。Gα可被位于质膜内侧的腺苷酸环化酶(AC)激活,后者催化ATP合成环磷酸腺苷(cAMP),使胞内cAMP水平上升,活化cAMP依赖的蛋白激酶A(PKA)。结合cAMP的PKA,其催化亚基与调节亚基分离,催化其他蛋白的Ser/Thr残基磷酸化,从而可进一步激活其他蛋白激酶。同时,β,γ亚单位可以在Ca2+协助下活化磷脂酶C(phospholipase C,PLC),分解磷脂酰肌醇三磷酸(PIP3)为二脂酰甘油(DAG)和肌醇三磷酸(IP3)。DAG可以激活PKC,进而引起其他蛋白磷酸化,而IP3可以激活Ca2+通道开放。已证实精子中存在G蛋白耦联受体。如GalT在结合ZP3糖链后,可通过G蛋白进行信号传递。而孕酮作用引发的顶体反应,也是由G蛋白介导、激活下游的磷酸酯酶A2(PLA2)和PLC来完成。另外,在精子中还发现去甲麻黄碱类物质(phenylpropanolamines)也是通过G蛋白作用调节cAMP浓度,加速精子获能,并抑制精子自发的顶体反应。在体细胞中,PKA可以通过MAPK途径或者ERK途径调节基因表达。但在精子中,PKA更多是激活PKC、蛋白Tyr激酶(PTK)或者是活化离子通道,以提高胞内磷酸化水平和Ca2+浓度。对哺乳动物精子来说,PLC调控的非选择性阳离子通道在Ca2+内流并诱导精子顶体反应的过程中起着重要作用。
受体酪氨酸激酶(RTK)途径
RTK是一种酪氨酸蛋白激酶,同时又担当受体的作用。其大致分为三部分:胞外部分为配体结合区,能与相应配体结合;由一段疏水氨基酸组成单次跨膜区;胞质侧是激酶活性区,具有多个酪氨酸残基。RTK介导的信号通路非常广泛,涉及细胞生长、分化、增殖以及代谢的调节。ZRK/hu9就是存在于精子膜上的RTK。当其与配体结合后,蛋白构象发生变化,相邻受体分子聚集,发生自我磷酸化(autophosphorylation),使得胞质侧激酶活性区的酪氨酸残基磷酸化。这不仅激活RTK的酪氨酸激酶活性,同时还为其靶蛋白提供停泊位点(docking site)。因为磷酸化的酪氨酸残基是SH2结合位点,能与含有SH2结构域的蛋白质结合并使之活化。这些含有SH2结构域的蛋白一般多为参与信号传递的酶蛋白或者是接头蛋白(adaptor protein),能够直接发挥效应或者将信号传递给其他信号蛋白,如hu9活化的下游信号效应蛋白主要是PLC和膜离子通道。
其他可能的信号传导途径
除上述外,还发现一些神经递质,如γ-氨基丁酸(gamma-aminobutyric acid,GABA)、乙酰胆碱(Ach)、甘氨酸,都可以引发顶体反应,但其生理学意义还不清楚。在精子表面已发现GABA受体、Ach受体和甘氨酸受体。GABA受体和甘氨酸受体是Cl-通道,通过Cl-外流可以引起膜电位去极化。而Ach受体是阳离子通道,可以促使Na+或者Ca2+内流引起膜电位去极化。因此推测其可能通过改变膜电位,激活电压门控的Ca2+通道,参与引起初次Ca2+波。如对小鼠甘氨酸受体和Ach受体进行人工突变的实验发现,精子的顶体反应发生率和体外受精率都有所下降。
