
过去数十年间,唾液腺被广泛用作研究外分泌器官功能的模型,这是因为唾液腺含有不同类型的细胞、受交感和副交感神经的控制、对多种神经介质有良好反应。唾液腺也被广泛用作一种研究外分泌腺发育的良好模型,因为唾液腺在出生时并无分泌功能,几乎全部功能是生后发育成熟的。许多外分泌腺在出生时已有全部或部分功能。然而,迄今为止对唾液腺的增龄性变化的研究相对较少,尤其是功能的改变,仍有待于大量探索。
唾液腺的形态和功能发育:唾液腺的功能发育始于出生后,这与其形态发育的特点有关。唾液腺的形态发育请参阅本书第一章。唾液腺在出生时已有良好的导管结构,有小叶分支,有末端胚芽,但不具备分泌终端细胞。这是出生时没有分泌功能的组织细胞学基础。出生时唾液腺已有闰管系统。已知闰管细胞是唾液腺发育的干细胞,它可向两个方向发育。向导管方向发育即分化为纹管细胞或颗粒导管细胞(后者仅限于啮齿类动物),向分泌终端方向发育即分化为末端小管细胞。出生时,末端小管主要由末端小管细胞和原腺泡细胞构成。生后1周内,原腺泡细胞经有丝分裂而发育为腺泡细胞,因而在1周末,细胞芽主要由腺泡细胞构成。原腺泡细胞在生后1~2周内耗竭,而末端小管细胞成为腺泡细胞的前身细胞。到第6周时,腺泡形成完毕,末端小管细胞消失。腺泡细胞无论来自哪种细胞,即原腺泡细胞或末端小管细胞,均可反复分裂,但增殖速度随年龄的增长而迅速减慢。唾液腺功能的发育是与上述细胞分化过程密切相关的,换言之,形态发育是功能发育的基础,但功能发育也可促进形态发育。刺激生后早期的动物的交感神经可加速发育过程就是典型的例子。
分泌功能的发育
如上所述,唾液腺分泌唾液的功能取决于分泌终端细胞的存在以及细胞分泌机制的成熟。以大鼠下颌下腺为例,出生后第1天时,平均腺体重量为15. 4mg,无唾液分泌功能。生后7天时达21mg,仍无分泌功能。直到生后14天时,才能观察到小量唾液分泌,其量只相当于成年分泌水平的3%,而腺体重量已达35. 6mg,相当于成年腺体的22%。生后3周(第21天)时,腺体重量达61. 9mg,相当于成年水平的38%,而唾液分泌量只有成年水平的12%。从图3-3可以看出,唾液分泌功能的发育明显慢于腺体重量的发育。而且,分泌终端细胞在出生后两周内已大量增殖,但仍然不能引起水与电解质的分泌反应,提示唾液分泌的机制或调节系统仍不成熟。
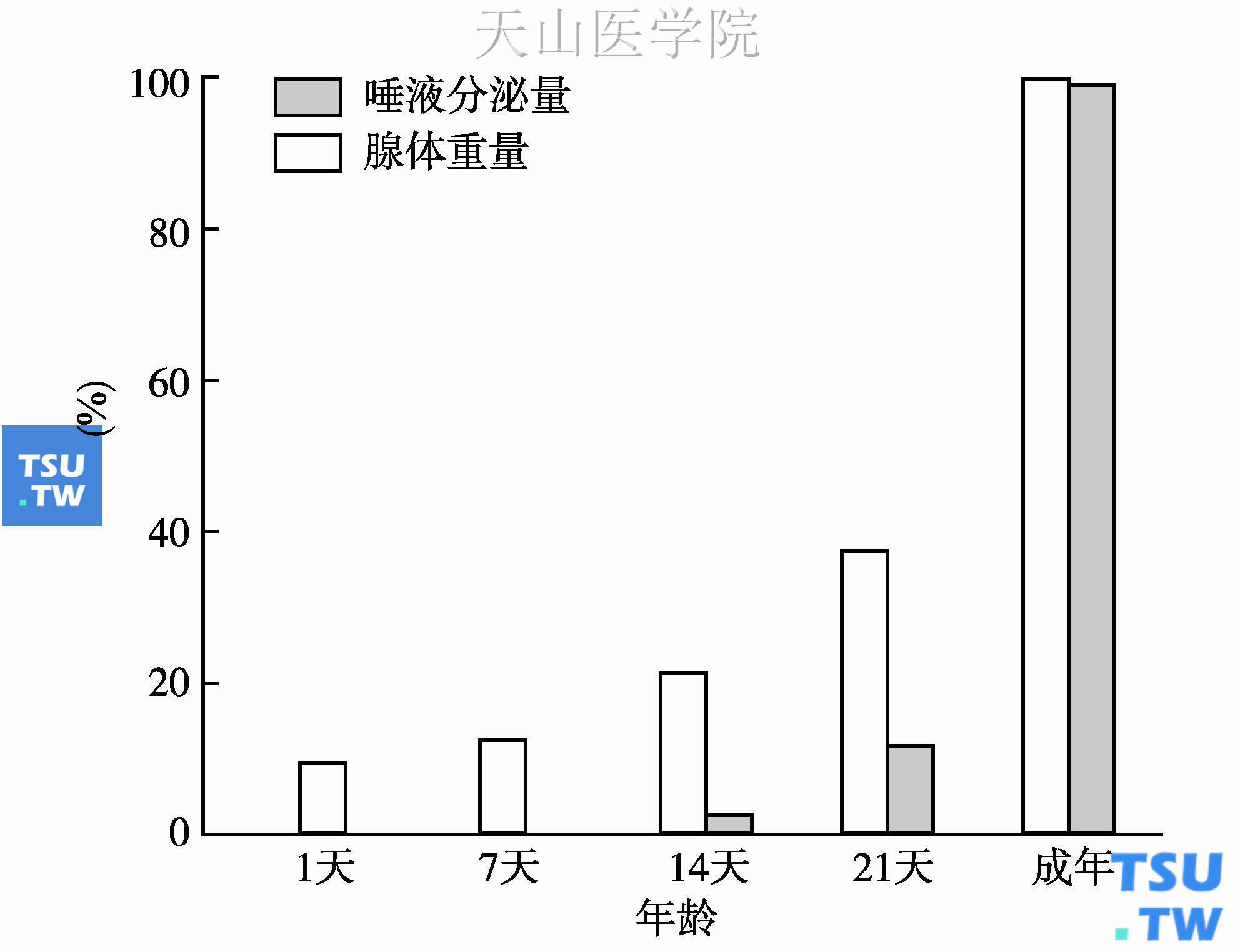
图3-3 大鼠下颌下腺腺体重量和唾液分泌量的发育性改变
腺体重量和唾液分泌量均以成年大鼠的数值为100%。唾液分泌是由腹腔内注射氨甲酰胆碱引起的。成年下颌下腺重量为162. 3mg;唾液分泌量为958. 6mg/h
蛋白质分泌功能的发育
严格地讲,关于蛋白质分泌功能的发育研究很少。多数研究集中在细胞内蛋白质含量方面。虽然细胞内蛋白质的含量并不能完全代表蛋白质的分泌,但在一定程度上亦可用其来推测蛋白质的分泌功能。事实上,胞吐作用及其调节系统亦有发育性改变。
(1)蛋白质含量和分泌:唾液腺细胞内蛋白质含量在出生后迅速增加,但不同腺体及不同蛋白质有自身的模式。
1)唾液淀粉酶:大鼠腮腺细胞内唾液淀粉酶含量在出生后的5天内增长100倍,第8天到第56天中增长1000倍,但用肾上腺素或异丙肾上腺素(40μmol/L)刺激第8天的腮腺仍无法引起淀粉酶的分泌。此后发育很快,到生后第15天,淀粉酶的分泌与25天及8周的大鼠已无差异。
大鼠下颌下腺淀粉酶含量的变化与腮腺不大相同。出生时与腮腺相当,在生后4天内继续增长达到最高水平,其后直线下降,到第15天时减到最低水平,此后又开始增加,到30天时达到15天时的两倍,此后再次减低,直到成年水平。
2) RNA酶和DNA酶:已知这些酶均大量存在于腮腺组织,而下颌下腺组织只含有微量。例如,DNA酶在腮腺中比在下颌下腺中高81 000倍,而RNA酶高540倍。在生前发育过程中,这两种酶均呈直线增长,到出生时已达很高水平。生后,腮腺内活性继续增长,直到30天时接近成年水平。而下颌下腺中这些酶在生后4天时达高峰,此后迅速减低,到50天以后达成年水平。
3)表皮生长因子(EGF):EGF的合成已被广泛研究。小鼠生后14天以前其下颌下腺颗粒导管细胞内并无EGF检出,但到第20天时则可检出相当含量。一般来讲,在第18天到20天之间可检出EGF多肽和mRNA。
4)神经生长因子(NGF):已经证明,NGF在小鼠下颌下腺颗粒导管细胞中的含量与颗粒导管细胞的发育平行。免疫组化研究证明,小鼠生后第3周时可检出NGF。在3~12周之间逐渐增加。
(2)加速蛋白质合成的因素:目前对可加速唾液腺蛋白质合成和分泌的因素了解很少。多数这方面的研究使用成年动物,因而无法肯定这结果也适用于发育中的动物。已知有几种因素可改变唾液腺细胞的发育和增殖,例如激素和神经介质。自主神经的交感或副交感系统均可调节唾液腺细胞的分化和生长。早期的研究多次证实,阻断交感或副交感神经可使生后早期的动物唾液腺生长迟缓,而早期给予自主神经激动剂如卡巴胆碱或异丙肾上腺素则可引起唾液腺细胞生长加快,腺体增大。然而,是否这些处理可使唾液腺细胞中分泌性蛋白质的含量增加、分泌时间提前,尚不清楚。唯一证明的作用是用甲状腺素处理生后第2周的小鼠可使NGF合成量增加,但这种处理不能改变生后第1周的小鼠的反应。有趣的是,成年小鼠NGF合成受性激素的影响,而生后前两周的动物对睾酮并无反应。这可能反映了唾液腺细胞尚无有效雄激素受体。
水与电解质功能的发育性改变
生后两周内的大鼠下颌下腺并无唾液分泌,这表明唾液腺尚不能分泌水和电解质。对大鼠下颌下腺的离子转运体系进行研究发现,这种水与电解质分泌的缺乏是由于离子转运体系的不成熟及调节这些转运体系的信号系统的不成熟。
迄今为止,对唾液腺离子转运体系的发育研究几乎只限于大鼠下颌下腺。
(1) Na+/K+泵:Na+/K+泵是唾液腺细胞水与电解质分泌及重吸收的能量来源,因此其生后发育性改变首先受到注意。Martinez等观察到,用特异性Na+/K+泵抑制剂乌本苷抑制生后1、7、14、21天的大鼠下颌下腺活组织切片引起了K+释放,表明钠泵出生后早期已有功能。K+释放量在发育的不同时期并无明显差异。生后1、7、14、21天的下颌下腺切片分别释放了28. 4%、21%、21. 1%、28. 2%K+。由于K+摄取是一种逆浓度梯度的主动转运过程,主要依赖Na+/K+泵活性和Na+-K+-2Cl-协同转运机制,因而Martinez等把大鼠下颌下腺切片培养在无K+溶液中15分钟,然后加入K+。唾液腺细胞摄取K+的量以生后1天的大鼠最高,为36%,生后7天的大鼠为12%,生后14天和21天的大鼠分别为8. 2%和1. 4%。这可能是由于在15分钟无K+溶液中,生后1天的大鼠释放的K+最多。这些结果清楚表明下颌下腺细胞在出生后早期已有很强的Na+/K+泵活性,但这些研究未能区别分泌终端细胞与导管细胞的Na+/K+泵活性。由于生后早期,唾液腺主要为导管系统构成,其Na+/K+泵活性可能反映了导管细胞Na+/K+泵,而生后3周时则反映了相当比重的分泌终端细胞的Na+/K+泵活性。测定细胞膜Na+-K+-ATP酶活性发现,该酶活性是逐渐发育增长的,生后1天时为6. 3μmol/(h•mg) DNA,而7、14、21天时分别为8. 4、9. 0、11. 7μmol/(h•mg) DNA。21天时已达成年水平。用分离的下颌下腺细胞进行K+转运测定,发现Na+/K+泵活性在非刺激状态下和毒蕈碱受体受到激动时,均表现出随年龄增长而增大,在生后第14天时达成熟水平。
(2) Na+-K+-2Cl-协同转运机制:新生大鼠的Na+-K+-2Cl-协同转运机制也已有部分功能,但非刺激状态下活性较弱。而且,Na+-K+-2Cl-协同转运机制所引起的K+摄取在非刺激状态下并不显示随年龄增长而变化(0~0. 8%)。但受到卡巴胆碱激动时,Na+-K+-2Cl-协同转运所摄取的K+大为增高,并显示出明显年龄差异。
(3) Cl-通道:Cl-通道在电解质分泌过程中起关键作用。用分离的大鼠下颌下腺细胞进行36Cl-外流研究,发现Cl-外流有发育性改变。当用0. 1μmol/L乙酰胆碱刺激细胞时,生后1天的大鼠下颌下腺细胞未发现Cl-外流,而生后7天的大鼠下颌下腺细胞有12%Cl-外流(表3-15)。从表中可以看出,Cl-外流随年龄增长而增大。应用细胞膜片钳术测定Cl-通道电流发现,生后2~3周内的大鼠下颌下腺细胞没有Cl-电导。用乙酰胆碱或肌霉素均未能引起Cl-外流。应用X线显微分析技术也证明,生后早期的大鼠下颌下腺细胞不能分泌Cl-。
表3-15 大鼠下颌下腺细胞Cl-外流的发育性变化

(4) K+通道:与Cl-通道的发育性变化类似,K+通道亦表现出发育性改变。用0. 1μmol/L乙酰胆碱激动毒蕈碱受体时,生后1天和7天的大鼠颌下腺细胞未能释放K+(表3-16),表明生后1周内,K+通道功能尚未成熟,但当用大剂量乙酰胆碱刺激时则可引起K+外流。
表3-16 大鼠下颌下腺细胞K+外流的发育性变化

综上所述,唾液腺细胞离子转运功能有一个生后发育过程。一般来讲,生后14天内Na+/K+泵和Na+-K+-2Cl-协同转运功能已获相当水平的发育,但尚未达成熟水平;而Cl-通道和K+通道尚未明显发育,推测生理性刺激不足以引起电解质转运。这与生后14天内无唾液分泌的观察是一致的。迄今,尚无有关其他离子和水转运机制,例如Na+-H+交换、Cl--HCO3-交换及水通道的活性的研究。
信号系统的发育
唾液腺蛋白质和水与电解质的分泌处于神经系统的严密控制之下。生后早期分泌功能的缺如或低下一方面可能是由于分泌性蛋白质尚未充分合成,分泌过程所需的机制仍不健全,如胞吐作用所需的蛋白质、离子转运过程所需的转运机制仍未发育成熟,另一方面可能是由于细胞信号传递系统尚未发育成熟。
细胞表面受体的发育性改变
现已清楚,自主神经受体即α1、α2、β肾上腺素能受体和胆碱能受体均经历生后发育过程,但每种受体的发育性变化不完全相同。迄今为止的绝大部分研究结果均来自于大鼠的下颌下腺细胞。
(1)胆碱能受体:大鼠下颌下腺细胞膜毒蕈碱型胆碱能受体在出生后1天时就已有中等密度,此后逐渐增长,到3周时达高峰。生后1~3周时受体密度高于成年大鼠,但其亲和力则明显低于成年大鼠,显示出受体蛋白质的不成熟。
(2)α1肾上腺素能受体:大鼠下颌下腺细胞α1肾上腺素能受体的发育性改变与毒蕈碱受体不同,出生时密度很低,以后逐渐增长,到3周时已接近成熟水平。受体亲和力也呈逐渐增长趋势,到3周时已稍微超过成年水平。
(3)α2肾上腺素能受体:α2肾上腺素能受体的变化趋势与α1受体的变化相反。出生时大鼠下颌下腺细胞已有很高密度的α2受体。出生后继续增高,到生后两周时达高峰。此后迅速减低,到6周时已达微量,成熟大鼠一般不能检出α2受体。与之相反,α2受体的亲和力却没有明显发育性变化,从出生时到成年,均在较稳定的水平。根据这些结果,α2肾上腺素能受体可能在唾液腺的发育过程中起重要作用。
(4)β肾上腺素能受体:β受体在出生时已有一定密度,此后逐渐增加,到第2周时达成熟水平。受体亲和力在出生时已经很高,此后稍有增长,到第3周时达成熟水平。
(5) P2核苷酸受体:迄今尚未见到系统性研究,但在大鼠下颌下腺细胞,刺激P2Y2受体引起的Ca2+动员以生后早期的反应最大,成年大鼠细胞的反应明显减低,提示存在发育性改变。
细胞内信号系统的发育性变化
大量研究表明,控制唾液腺蛋白质分泌以及水与电解质转运的信号系统,即cAMP-PKA和磷酸肌醇-Ca2+系统,均有发育性变化。显然,这些信号系统的成熟对于唾液分泌有重要意义。
一、cAMP-PKA系统
大鼠出生后早期,腮腺细胞内没有cAMP生成,显然这是由于腺苷酸环化酶尚未成熟。用40μmol/L肾上腺素或异丙肾上腺素刺激生后8天以前的大鼠腮腺细胞的α1和β受体,不能引起cAMP产生,到15天时则可;到25天时cAMP产生增长8倍,56天时增长8~12倍。与此稍有不同,大鼠下颌下腺细胞腺苷酸环化酶的活性在出生时已经较高,生后没有明显变化。对此,仍有争议。有的研究认为,生后14~28天时,腺苷酸环化酶活性增加一倍。这种腺体间差异的意义仍不清楚。迄今尚未见到有关PKA的发育性变化的报道。
二、磷酸肌醇-Ca2+系统
尽管生后早期的大鼠下颌下腺细胞并不能分泌水与电解质,但IP3产生和Ca2+动员反应却明显大于成熟细胞。这种过度反应随年龄的增长而减小,一般于5周之后与成年大鼠无明显差异。测定细胞内Ca2+池摄取和释放45Ca2+发现,生后早期的细胞低于成熟细胞。不成熟的动物这种IP3的过度生成及Ca2+动员反应的机制尚不明了。
1) G蛋白:由于毒蕈碱受体结合时首先激活三聚G蛋白,因此对其功能进行了测定。生后早期的大鼠下颌下腺细胞表达Gqα蛋白少于成年腺体细胞。而且幼鼠下颌下腺细胞Gqα与毒蕈碱受体的亲和力也明显低于成年大鼠细胞。这些结果表明,IP3生成反应过高并不是由于Gqα的过度表达。
2)磷脂酶C:年幼大鼠下颌下腺表达的磷脂酶C亚型明显高于成年大鼠。其中磷脂酶Cβ1亚型在出生后1天时为成年水平的2. 9倍,第7天时为2. 1倍,第14天时为1. 3倍。而磷脂酶Cβ3亚型出生后第1天为成年水平的48倍,第7天时减低为14. 3倍,第14天时为5. 9倍;磷脂酶Cγ1亚型生后第1天为成年水平的3. 7倍,此后继续增高,到第7天时为8. 3倍,第14天时仍为4. 3倍。磷脂酶Cγ2亚型出生后第1天为成年的1. 8倍,此后增长为第7天时的2. 6倍及第14天时的3. 2倍。从这些结果可以看出,磷脂酶Cβ3、γ1 和γ2亚型在生后第14天时仍然极不成熟,仍需时间继续发育。显然,幼鼠下颌下腺细胞IP3反应过度的原因在于磷脂酶C的过度表达。
3) Ca2+池:未成熟大鼠下颌下腺细胞内Ca2+池的容量较小。经Ca2+泵转运进入Ca2+池的Ca2+亦少于成年大鼠。用内质网Ca2+泵抑制剂毒胡萝卜素处理引起的Ca2+释放在幼鼠细胞也小于成年细胞。这些结果均证明,幼鼠细胞内Ca2+池小于成年大鼠细胞,因而毒蕈碱受体激动时所引起的过度Ca2+增高不是由于Ca2+释放量高于成年大鼠细胞。
4)内质网Ca2+泵:胞质内游离Ca2+的减低依赖两条途径,即内质网和质膜Ca2+泵将Ca2+泵入内质网或泵出细胞。研究发现,幼鼠下颌下腺内质网Ca2+泵活性低于成年大鼠,因而转运入Ca2+池的Ca2+明显低于成年大鼠细胞。
5)质膜Ca2+泵:将Ca2+泵出细胞是由质膜Ca2+泵所驱动的。毒蕈碱受体激动可增强质膜Ca2+泵的活性。幼鼠下颌下腺细胞Ca2+外流明显小于成年大鼠细胞,无论非激动或激动状态均如此,表明幼鼠细胞Ca2+动员反应大于成年细胞至少部分是由于Ca2+泵尚未成熟,不能将胞质中的Ca2+迅速转运到细胞外。
三、发育过程中信号系统之间的相互作用
已知PKA系统和PKC系统对生长发育过程有影响。在许多细胞的生长分化过程中,PKA和PKC的活化可调节早期反应基因及转录因子如c-fos,c-jun的表达,也调节核糖体蛋白如S6的表达。这些激酶系统和基因表达可调节细胞分裂周期,其作用机制是通过活化其他激酶系统及磷酸化网络,例如分裂原激活的蛋白激酶系统及p70 S6激酶系统。
1) cAMP-PKA系统与磷酸肌醇信号系统的相互作用:激活成熟大鼠下颌下腺分泌终端细胞PKA可明显抑制乙酰胆碱所引起的Ca2+释放反应。用细胞膜通透的cAMP异构体二丁基cAMP处理使Ca2+反应减低50%。同样用腺苷酸环化酶激活剂毛喉素处理亦可使Ca2+反应减小52%。与之不同,用相同的物质处理从生后1天的大鼠下颌下腺分离的细胞并不影响乙酰胆碱引起的Ca2+的反应。例如用二丁基cAMP处理生后1天的下颌下腺细胞后,乙酰胆碱所引起的胞质Ca2+浓度增加为对照的108%,并无显著意义。发生这种现象的机制尚不确定,但极有可能是由于PKA系统尚未成熟,或者磷酸肌醇-Ca2+信号系统对磷酸化作用尚不敏感。
2) PKC与磷酸肌醇-Ca2+信号系统之间的相互作用:PKC可能在调节内质网Ca2+泵活性方面起重要作用。激活成年大鼠下颌下腺分泌终端细胞PKC可抑制毒胡萝卜素所引起的Ca2+释放,而抑制PKC则无作用,机制不明。推测PKC可能增强Ca2+泵活性或减低Ca2+泵对毒胡萝卜素的敏感性。生后1天的大鼠下颌下腺细胞则表现为不同的反应。激活PKC对毒胡萝卜素引起Ca2+的释放无影响,而长时间(30分钟)激活则可加强Ca2+释放反应。
迄今为止,其他信号传递系统与上述系统的相互作用还未被广泛研究,尤其是发育性变化,知之甚少。然而,阐明这些问题会促进对唾液腺发育过程的了解。
