
基本内容
1971年Eversole在观察唾液腺胚胎发育的基础上首次提出了正常唾液腺中存在储备细胞的概念。他在观察腭部小唾液腺的胚胎发育过程中发现,口腔表面上皮细胞向深部生长分化为双层导管细胞,且内层或腔面层细胞来源于外层或基底层细胞。由于基底细胞能够产生导管腔面细胞和腺泡细胞,发挥着干细胞的功能,故被认为是储备细胞。随着唾液腺的发育成熟,这些细胞留存于排泄管的基底层,排泄管基底细胞负责排泄管的腔面及基底细胞的更新,同时它也是闰管细胞的前体细胞,这些闰管前体细胞负责闰管、纹管、腺泡的发育、更新,因而排泄管基底细胞和闰管细胞均为多能的储备细胞,此即唾液腺的半多能双储备细胞理论。
至1977年,Regezi和Batsakis采用了唾液腺储备细胞的概念,并在此基础上加以补充和发展。他们提出腺泡细胞为高度分化的细胞,纹管细胞也是分化较好的细胞,而闰管细胞是相对未分化的细胞,肌上皮细胞位于腺泡及闰管基底部具有收缩功能。排泄管储备细胞分化出排泄管的柱状及鳞状细胞,闰管储备细胞除分化为腺泡细胞、闰管细胞和纹管细胞外,还分化出肌上皮细胞(图5-1)。
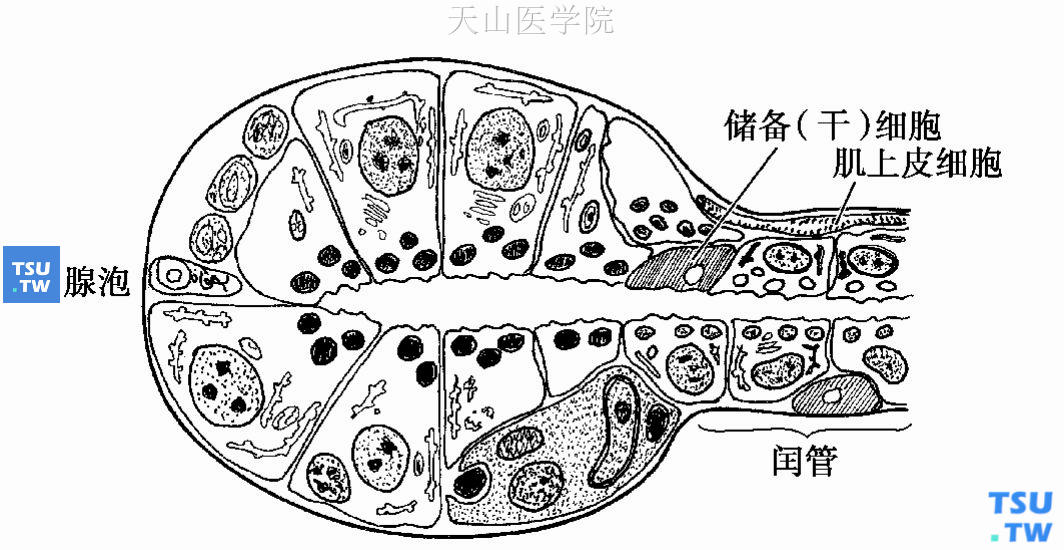
图5-1 储备细胞是多潜能的细胞,能够分化为导管细胞、腺泡细胞和肌上皮细胞
在储备细胞概念的基础上,Regezi和Batsakis提出了唾液腺肿瘤的组织发生机制。他们指出唾液腺肿瘤的发生不是来源于成熟细胞的去分化,而是来源于排泄管储备细胞和闰管储备细胞这两种未分化储备细胞,这就是肿瘤起源的双细胞理论(bicelluar theory of origin)。唾液腺的鳞状细胞癌、黏液表皮样癌、乳头状黏液腺癌来源于排泄管储备细胞,而多形性腺瘤、单形性腺瘤、肌上皮瘤、嗜酸细胞腺瘤、腺样囊性癌、腺泡细胞癌、腺癌来源于闰管储备细胞。虽然以后不断有研究表明,许多唾液腺肿瘤的组织来源远非如此简单,并且某些肿瘤的来源细胞也并非如Regezi和Batsakis所述(有关内容详见第四节),但肿瘤起源的双细胞理论对于研究唾液腺肿瘤的组织发生起了重要的推动作用。
以后,Batsakis等(1980,1983)又对唾液腺肿瘤的发生机制作出了进一步解释。他们认为,唾液腺的发生来源于表面口腔上皮,其各部分的形态生成及细胞分化如图5-2所示:
半多能双储备细胞理论认为,唾液腺肿瘤的形成是由于在唾液腺组织更新或再生过程中,干细胞或储备细胞的肿瘤基因受到刺激而激活,使组织在形态生成及细胞分化时细胞发生了异化所致。
正常腺体的细胞增殖
一般认为,只有能够进入细胞周期进行增殖的细胞才有可能发生肿瘤。腺体组织内新细胞的生成至少可来源于下列三种方式之一:①复制,即现有细胞的有丝分裂;②分化,例如从闰管细胞分化出特定的腺泡细胞;③转化,指从一种已分化细胞到另一种具有不同遗传表型的细胞。其中转化方式由于表明的是一种已经具备其最终表型的高度特化的分化细胞可改变其形态、功能机制而成为另一种类型的分化细胞,因而此种方式发生的可能性最小。转化方式中其实还包含了去分化的概念,而Batsakis等认为去分化是不可能发生的。
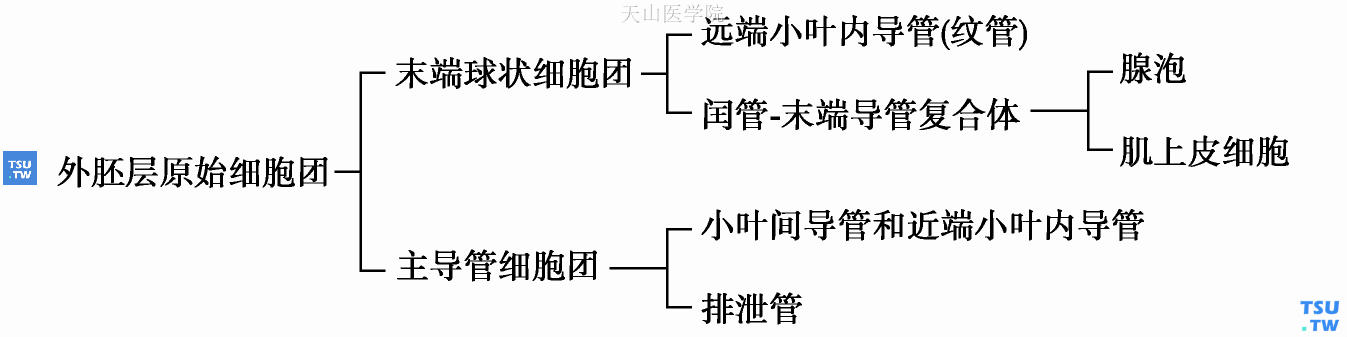
图5-2 唾液腺组织形态生成及细胞分化方式
在胚胎期和新生儿腺体中的腺泡细胞存在着复制现象,至此之后细胞的复制急剧减少。Batsakis等曾引用一些腺体导管结扎后、激素诱导的胰腺炎、乙基硫氨酸导致的胰腺坏死等实验说明腺体中增殖活性最高的是导管或末端导管细胞,这其中包括Walker and Gobe的实验,此实验表明,在唾液腺导管结扎或部分切除后新生腺泡细胞来源于末端导管或闰管,因而Batsakis等认为,实验中腺泡和闰管的标记指数表明,在腺泡细胞的新生过程中,腺泡细胞是由末端导管或闰管分化而来,即上述三种方式中的分化方式,而另外两种方式即腺泡细胞的自我复制以及由其他细胞转化而来的方式并不存在。但以后有学者证实,腺泡细胞是有复制能力的,虽然它的复制率较低,关于此点详见后述。
肌上皮细胞与唾液腺肿瘤发生
肌上皮细胞位于腺上皮细胞和基底膜之间,具有收缩功能。虽然电镜下见,其细胞结构特点与平滑肌细胞有某些相似,如存在肌微丝、密斑等结构,但形态学观察及免疫组化均证实细胞内还含有中间丝细胞角蛋白,而后者一般只在上皮细胞中存在。且从部位上来看,肌上皮细胞位于近腺上皮的基底膜内侧,也提示其为上皮性来源。由此可见,肌上皮细胞兼具上皮和间叶细胞的特性,但细胞为上皮性来源。大量实验证明,肌上皮细胞来源于外胚层上皮。
半多能双储备细胞理论特别重视肌上皮细胞在唾液腺肿瘤发生中的作用,并认为唾液腺中肌上皮细胞的来源可能有两种方式,见图5-3:
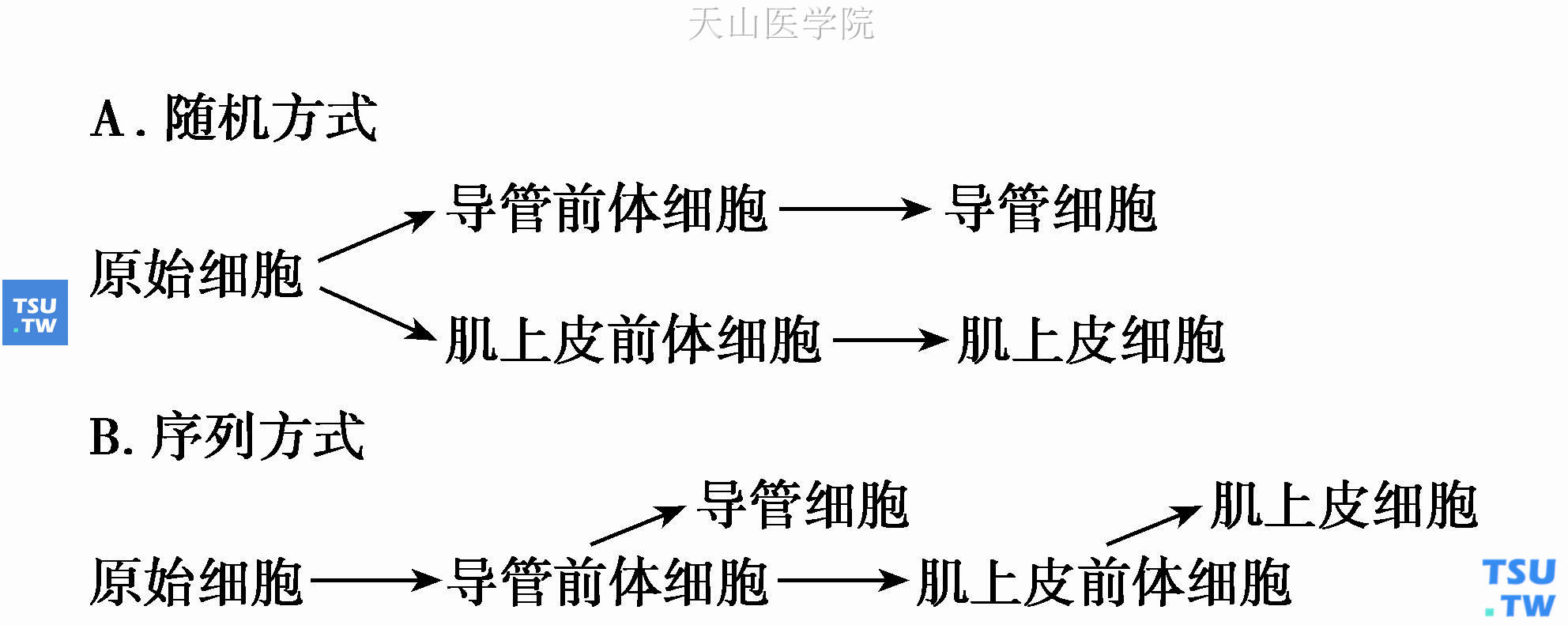
图5-3 形成肌上皮细胞的两种可能方式
第一种方式中,导管细胞和肌上皮细胞由不同的前体细胞分化而来,而在第二种方式中,两前体细胞本身就来自同一原始细胞系,是原始细胞顺序分化的结果,原始细胞实际上即是具备多潜能分化的干细胞,而导管前体细胞相当于闰管储备细胞。现在认为肌上皮细胞的来源更趋向于第二种方式。如Shirasuna等的实验证明,在体外培养的闰管样克隆化细胞中可分化出肌上皮细胞,提示后者来自闰管样细胞。
Batsakis等认为肌上皮细胞在唾液腺肿瘤发生中扮演了重要的角色。同为外分泌腺,胰腺组织由于缺乏肌上皮细胞,因而其发生的肿瘤远未像唾液腺肿瘤这样具有多样性。出生后肌上皮细胞的更新来源于闰管储备细胞。虽然正常腺体中肌上皮细胞的增生能力是很低的,但唾液腺肿瘤中的肿瘤性肌上皮细胞则不同,有着较强的增殖能力。肿瘤性肌上皮在形态上的表现具有双重性,可表现为间质型的成纤维细胞样、黏液样,以及上皮型的浆细胞样、上皮样。肿瘤性肌上皮一方面细胞自身构成了唾液腺肿瘤的一部分,在某些肿瘤中可以见到大量肌上皮细胞的排列,如多形性腺瘤、肌上皮瘤、低度恶性多形性腺癌等;另一方面它和肿瘤基质的形成密切相关,例如它可形成多形性腺瘤中的间质成分和腺样囊性癌中的基底膜样物质。
从生物学观点来看,出现肌上皮样细胞的唾液腺肿瘤其转移率低、生存期长因而恶性程度低。实验证明有肌上皮样细胞存在的细胞系,不论是在体内还是体外其恶性程度均低得多。此点在乳腺癌中也得到证实。在不同类型的唾液腺肿瘤中,高度恶性的肿瘤,如唾液腺导管癌和高度恶性黏液表皮样癌无肌上皮样细胞,而有肌上皮样细胞的肿瘤如低度恶性多形性腺癌和上皮-肌上皮癌则为低度恶性肿瘤。
干细胞与唾液腺肿瘤发生
半多能双储备细胞理论认为肿瘤的发生是由于细胞在发生和再生过程中发生了异化所致,是干细胞发生突变的结果。突变导致细胞分化过程的中断,随之产生一些基因表达改变的细胞,这种基因改变是朝有利于恶性细胞生成的方向发生。对于唾液腺肿瘤来说,因此而产生的细胞更易失去其分化为肌上皮细胞的能力。干细胞或储备细胞为唾液腺再生细胞的来源,其肿瘤基因容易受到刺激而激活。这些细胞一方面可分化为肌上皮细胞,另一方面可分化为腺上皮细胞。
传统的肿瘤分类是以肿瘤细胞与正常组织细胞的形态相似性为依据,但用此来解释唾液腺肿瘤的发生时有两点困难,一是唾液腺肿瘤的异质性(heterogeneity),即常见肿瘤中存在间质样分化,二是照此理论推理,一些分化成熟的细胞如腺泡和纹管细胞在肿瘤形成时一定要发生去分化,这在Batsakis看来几乎不可能。而半多能双储备细胞理论认为用储备细胞或干细胞理论能很好地解释肿瘤的形成机制。他们认为在唾液腺的发育过程中存在着三个阶段:未分化阶段、初分化(protodifferentiated)阶段和分化成熟阶段。在唾液腺形成和细胞分化的不同阶段有不同类型肿瘤的形成。如在唾液腺发育的未分化阶段形成未分化癌,初分化阶段形成多形性腺瘤、基底细胞腺瘤、腺样囊性癌等。
由于干细胞或储备细胞可分化为肌上皮细胞和腺上皮细胞,这种细胞形式的转变可用来解释不同唾液腺肿瘤组织病理类型间的既相互类似又相互区别,同时也避免了各唾液腺肿瘤来自不同细胞的观点。确实,各类唾液腺肿瘤并不是相互毫无关联的,在以某种类型为优势的肿瘤中,常可见到其他类型的肿瘤,此特征进一步支持了干细胞理论,并提示肿瘤的进展与干细胞及其子代细胞对分化信号的反应异常有关。
综上所述,半多能双储备细胞理论认为,闰管细胞和排泄管基底细胞是唾液腺的储备细胞或干细胞,在唾液腺再生和肿瘤形成中起着重要的作用。
半多能双储备细胞理论在20世纪70~80年代被大家广泛接受,并被作为唾液腺肿瘤分类的基础。但是不论在正常、损伤或再生的唾液腺中,除了闰管细胞和排泄管基底细胞外,还可看到其他一些增生细胞,这些细胞的增殖能力如何?有何意义?半多能双储备细胞理论尚未给出明确的回答。
近年来,有学者对唾液腺恶性肿瘤的研究发现,被认为来源于排泄管储备细胞的肿瘤和来源于闰管储备细胞的肿瘤在蛋白表达上有显著不同,来源于排泄管储备细胞的唾液腺导管癌、黏液表皮样癌、非特异性腺癌环氧化酶-2(Cox-2)、S期激酶相关蛋白2(S-phase kinase associated protein 2,Skp-2)高表达,而来源于闰管储备细胞的腺泡细胞癌、基底细胞腺癌、腺样囊性癌、多形性低度恶性腺癌等Cox-2不表达或表达很低,但P27高表达,Skp-2低表达,研究结果支持唾液腺肿瘤发生的双细胞理论。
