
下丘脑(hypothalamus)是人体的神经-内分泌高级调节中枢,也是神经-内分泌调节的转换站。神经-内分泌研究创立了神经内分泌学(neuroendocrinology),其范围从最初控制垂体激素分泌的下丘脑激素扩展到了中枢神经系统调节内分泌功能的所有方面。
成年人的下丘脑重约4g(占全脑重量的1%以下),但在维持人体内环境稳定和神经-内分泌功能方面起着十分重要的作用,并与水电解质平衡、摄食、生殖、免疫、行为、心理和衰老等生命活动的关系十分密切。下丘脑神经细胞有分泌激素功能,这种分泌方式称为神经分泌(neurocrine),分泌的激素称为神经激素(neurohormone),分泌激素的细胞称为神经分泌细胞(neurosecretory cells)。
间脑内有第三脑室,并经室间孔与两侧脑室相通。间脑后部通向导水管,上部为松果腺,后下方与大脑脚相接。在大脑矢状切面上,可见第三脑室侧壁后方突起的丘脑,其下部为下丘脑。下丘脑的界限不甚分明,前为视交叉及终板,后为乳头体与脑脚间窝,上为大脑前联合及丘脑下沟,下丘脑向下伸展与垂体柄相连(下图)。
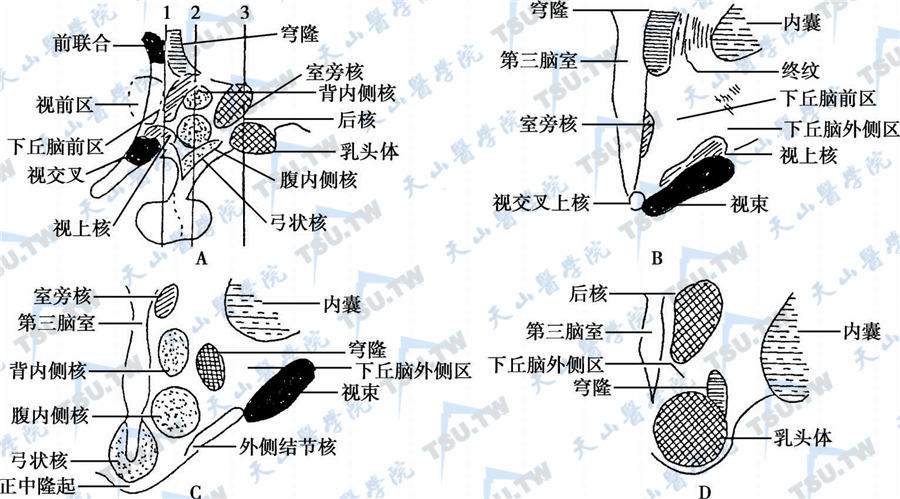
下丘脑的结构分区
注:A:矢状切面示意图;B、C、D:分别按A的1、2、3处的冠状切面示意图
正中隆突是下丘脑联系垂体的结构基础
下丘脑由前至后分为3个区:
- 前区(或视上区)位于视交叉之上,其前为居于前联合及视交叉之间的视前区;
- 中区(或结节区,灰结节)为下丘脑最宽处,与垂体相距最近,灰结节的中央部分称为正中隆突(median eminence),垂体柄(pituitary stalk)由此伸出,结节区外侧为下丘脑外侧区,内有大量神经纤维;
- 后区(乳头区)包括乳头体及其所含的神经细胞。
正中隆突和垂体的距离最近,关系最密切,是下丘脑调节垂体功能的最重要部位,也是各种促(抑)垂体激素必经的共同通道。有人认为,正中隆突亦是一种内分泌腺,因为:
- 此区域含大量神经末梢和神经分泌细胞,可合成和分泌神经调节肽(神经介素,neuromedin);
- 正中隆突的下端与垂体柄相连,处于灰结节的正中,相当于门脉系统第一微血管网所供应的区域,其内部包括大量神经纤维、神经胶质细胞(neuroglial cells)、门脉血管、脑室膜细胞(伸长细胞,tanycytes)和特殊分化了的神经胶质细胞,如垂体细胞(pituicytes);
- 正中隆突含腺垂体释放激素和释放抑制激素。
根据分泌颗粒的大小,可将神经轴突分为数种类型。不同的下丘脑激素位于不同类型的神经轴突中。不过,同一轴突中很可能含肽类和胺类递质。除正中隆突处,室管膜细胞(ependymal cells)的基底部突出,显示分泌活动并和血管系统相接。第三脑室周围的终板、联合下器、穹隆下器和第四脑室后区统称为“环脑室器”。
视上核和室旁核的神经激素沿轴突移行至神经末梢和血管相接处。血管加压素(AVP)调节血液渗透压、体液平衡和血压,而催产素在分娩时引起子宫收缩和乳汁分泌。下丘脑与腺垂体为神经-血管联系,下丘脑的神经轴突在正中隆突、垂体柄处与垂体门脉系的第一微血管丛相接,促(抑)垂体激素在此处释放入血,然后沿门脉血管到达腺垂体,兴奋(或抑制)腺垂体激素的分泌。不同的门脉血管引流下丘脑促垂体区的不同部位。通常情况下,每一支或一组门脉血管只含某一种释放激素和(或)释放抑制激素,供应腺垂体某一类型(分泌某一种腺垂体激素)细胞。近年发现,腺垂体含有大量神经纤维,中枢神经可能与腺垂体有直接神经纤维联系。
下丘脑神经核团按部位分区而视上核与室旁核功能特殊
下丘脑不同部位和不同核团的功能及形态并不相同,一般可分为以下几个功能区:
- 下丘脑前区:与促性腺激素的分泌有关,在雌激素的兴奋作用下(正反馈作用),引起月经中期促性腺激素(主要是LH)释放,促进排卵;
- 下丘脑中后区:也影响促性腺激素的分泌,其作用受雌激素的抑制(负反馈作用),此区域大约与促性腺激素的经常性分泌有关;
- 下丘脑前区和前腹室周核区:下丘脑前区与促甲状腺激素(TSH)的分泌有关;而前腹室周核区位于室周部之腹侧,紧接终板之后,为一密集而深染的卵圆形核团,此处亦含有许多促甲状腺激素释放激素(TRH)细胞,可调节TSH的分泌;
- 近正中隆突区:与生长激素(GH)的分泌有关;
- 促肾上腺皮质激素(ACTH)分泌区:控制ACTH分泌的区域较为广泛,因此下丘脑损害不容易使正中隆突的促肾上腺皮质素释放激素(CRH)浓度下降;
- 正中视前核区:位于第三脑室前沿,为一群密集的小细胞,是下丘脑渗透压敏感区之一;
- 视上核(suprachiasmatic nucleus)区:位于视交叉上方(细胞小而密集),是昼夜节律的产生部位;
- 室旁核区:为下丘脑大核团之一,位于下丘脑的室周部,含有1300~2000个神经元,特别是分泌催产素或AVP的神经元相对集中。下丘脑附近有若干渗透压敏感区,可感受到血浆渗透压1%的微弱变化。
- 前腹部的室旁核(anteroventral periventricular nucleus)和弓状核(arcuate nucleus)神经元表达kiss-1基因,其转录产物为kisspeptin;kisspeptin可激活G-蛋白耦联受体GPR54,在青春期发育的启动与下丘脑-垂体-性腺轴的功能调节中起了重要作用,而kisspeptin的靶细胞是正中隆突的促性腺激素(GnRH)神经元。因此,下丘脑是GnRH的脉冲起搏点(GnRH pulse generator),而KISS1R或神经激肽(neurokinin B,NKB)位于下丘脑,突变可引起低促性腺激素性性腺功能减退症。
神经分泌细胞产生神经激素/神经调节肽/类固醇激素/旁分泌激素
神经纤维起源于下丘脑底部、弓状核(漏斗核)、腹内侧核、室周核(periventricular nucleus)及视交叉上核,终止于正中隆突。
- 大神经分泌细胞:体积较大,位于视上核及室旁核,其轴突形成视上(室旁-垂体束,轴突末梢终止于神经垂体(垂体后叶)内,小部分终止于正中隆突。下丘脑的终板血管器(organum vasculosum lamina terminalis,OVLT)是调节AVP释放和饮水行为的主要部位。视上核主要产生AVP,而室旁核主要合成催产素(oxytocin),但此两个神经核团还可合成许多其他激素,神经分泌颗粒可被铬明矾-苏木精桃红选择性染色。神经分泌激素沿轴突下降,储存于神经垂体内。
- 小神经分泌细胞:产生多种调节垂体激素的神经肽(neuropeptides),包括促进或抑制腺垂体激素分泌的各种释放激素或释放抑制激素。神经纤维起源于下丘脑底部、弓状核(漏斗核)、腹内侧核、室周核及视交叉上核,终止于正中隆突。
- 神经胶质细胞和星形细胞:神经胶质细胞和星形细胞有重要功能。研究发现,哺乳动物的神经胶质细胞(neuroglial cells,gliocytes)能够合成各种类固醇激素,如孕酮(progesterone)、17-羟孕酮(17-hydroxyprogesterone)、睾酮(testosterone)、硫酸脱氢异雄酮(dehydroepiandrosterone sulfate)等,这些激素具有调节神经分泌、行为等功能。星形细胞(astrocyte)表达iNOS,而一氧化氮(NO)是兴奋性神经突触传递(excitatory synaptic transmission)的非典型信使物质。伸长细胞和垂体细胞与血管紧密相连。这些细胞通过调节与周围血管的“开放”和“关闭”来调控下丘脑激素的释放。因此,伸长细胞和垂体细胞具有血-脑脊液屏障功能,但垂体细胞还表达阿片肽、AVP受体和β-肾上腺素能受体,可接受多种激素或神经递质的调节;伸长细胞可能具有下丘脑神经轴突的“操纵功能”(guiding function),调节垂体-门脉系统的物质转运。
下丘脑神经细胞受多种因素调节
下丘脑神经细胞具有以下主要功能:
- 神经递质(transmitters)和神经调质(modulators)功能:神经激素的作用不是单一的,各种神经激素之间形成复杂的调节网络,并对下丘脑神经细胞本身以及它们所支配的靶细胞有明显调节作用;
- 信号整合功能:可将接受的多种信号整合为一种信号,并以某种神经激素为介导,作用于其他神经细胞或靶腺细胞;
- 靶细胞功能:许多神经分泌细胞膜或细胞质内含有多种激素受体,可接受循环血液或旁分泌而来的激素作用,并作出相应的激素分泌反应。
下丘脑激素的一般调节
目前发现,对垂体有直接调节作用的下丘脑激素除经典的TRH、GnRH、CRH、GH释放激素(GHRH)外,还有泌乳素(PRL)释放肽(PRL-releasing peptide,PRLrP)、生长抑素(somatostatin,SS)、血管活性肠肽(VIP)、血管加压素(vasopressin,AVP)、催产素和葛瑞林(ghrelin)。根据以上原因,人们提出了下丘脑-垂体单位(hypothalamic-pituitary unit)的概念。但是,垂体-门脉系统在转运下丘脑激素的分工方面存在种属差异,不同种属的下丘脑激素在分泌后向垂体的转运途径和方式并不相同,有的种属主要通过长门脉系统转运,而另一些种属主要由短门脉系统转运。
生理情况下,下丘脑神经细胞接受外周组织的神经体液调节。例如,颈动脉体(carotid body,CB)是一种外周化学感受器,颈动脉体的球状细胞与岩神经节神经末梢形成突触联系,主要接受低氧、高二氧化碳和酸中毒的刺激,并释放多种神经递质,其中兴奋神经中枢的递质有多巴胺、5-羟色胺和乙酰胆碱。此外,球状细胞还接受血管张力、温度和渗透压的刺激,并将这些物理化学信号转变为神经信号,反馈给下丘脑和其他神经中枢。
下丘脑激素的应激调节
在病理情况下,下丘脑神经细胞是应激反应的调节中枢。例如,重症患者T3下降,T4正常或低下,TSH降低或正常,提示应激可能通过细胞因子直接作用于垂体TSH细胞,使之对TRH的反应性降低。IL-1β可直接抑制垂体TSH的释放,TNFα和IL-1β可影响TRH诱导的TSH基础分泌。
kisspeptin及其受体调节性腺激素分泌和生殖功能
kisspeptin是kiss1基因的表达产物。1996年,首先发现kisspeptin是一种肿瘤转移抑制因子。kiss1中的“ss”代表抑制因子序列(suppressor sequence),又称为转移抑制素(metastin,含54个氨基酸残基),kisspeptin的前缀“kiss”是为了纪念该因子的发现地宾夕法尼亚的Hershey镇(巧克力品牌“Hershey’s Kisses”生产地)。研究肿瘤者喜欢使用metastin,而其他学者习惯称为kisspeptin。2001年鉴定了kisspeptin,其分子对RF胺(RFamide,Arg-Phe-NH2)呈高亲和性,相应的受体是G蛋白耦联膜受体(orphan G proteincoupled membrane receptor)。kisspeptin受体先后被称为AXOR12、hOT7T75、GPR54、KISS1R、KiSS1或metastin受体。Gottsch等建议统一使用kisspeptin和kisspeptin受体(KISS1R),并统一命名为“kiss1r”(大鼠)或“KISS1R”(人类)。
kiss1基因
kiss1基因的初级产物含145个氨基酸残基,裂解后的54氨基酸残基产物称为kisspeptin-54。顶端酶(vertases)是其降解酶,kisspeptin-54在体内的分解产物有kisspeptin-10、-13和-14,它们都含RF胺,而且与Kiss1r的亲和性相同。kiss1r跨膜7次,N端含3个糖化部位。在结构上,kiss1r与galanin受体相似(相同序列45%),但与galanin无结合活性。在筛选kiss1r激动剂的过程中,发现数种RF胺(Phe-Met-Arg-Phe-NH2)和RW胺神经肽可与Kiss1结合。kiss1r与kiss1导致磷脂酶C(PLC)活化,其信号通路与Gq/11关联,并进一步激活细胞内第二信使三磷酸肌醇(IP3)和二酰甘油(diacylglycerol,DAG)。这些信号分子再引起细胞内Ca2+释放与蛋白激酶活化,开放TRPC通道,刺激GnRH分泌,通过DAG和(或)Ca2+,抑制内向性(inwardly rectifying)钾通道。此外,kiss1r也刺激花生四烯酸释放,激活ERK1/2、p38和Rho,引起应激性纤维形成。IL2~IL10可引起Kiss1r结构重排,提示一些分子与kisspeptin的GPCR/G蛋白信号存在“对话”现象,而KISS1R的活化性突变(Arg386Pro)可引起性早熟。
kisspeptin蛋白及其受体
是调节生殖功能的关键因子。下丘脑的神经核团中散布有kiss1/kisspeptin神经元,这些神经元直接支配并刺激GnRH神经元,进而调节性腺功能。有些kisspeptin神经元也表达辅神经递质(cotransmitters),如强啡肽(dynorphin)、NKB、雌激素受体和雄激素受体等。不论性别,kisspeptin神经元都是性腺类固醇激素的靶细胞。kisspeptin调节GnRH的合成与分泌,并产生排卵前GnRH/LH促发分泌峰,启动青春期发育。此外,kisspeptin信号还是某些肿瘤转移、大脑、血管和胎盘功能的重要调节因子。
kisspeptin功能
kisspeptin具有多种生理作用,其中最主要的是调节生殖和性激素分泌,刺激GnRH分泌,在类固醇类型激素的正反馈和负反馈中均起了关键作用。大多数kiss1神经元表达雌激素受体和孕激素受体,大约40% 的kiss1神经元表达瘦素(leptin)受体,因而kisspeptin也可能是联系营养和生殖功能的物质基础,kiss1神经元受光周期(photoperiod)调节,并表达强啡肽A与神经激肽B(NKB),kiss1神经元含交感和非交感神经突触,提示kiss1神经元可同时接受兴奋性和抑制性信号,NKB激动剂抑制LH分泌,kisspeptin、dynorphin、NKB和谷氨酸都参与了GnRH脉冲性分泌的调节。分泌AVP的kiss1神经元可能接受视上核的传入神经信号。kisspeptin信号可直接调节GnRH,刺激GnRH分泌。此外,kisspeptin也通过中介神经元(GABA神经元)引起GnRH分泌。
AVP神经元是一种两性神经元(dimorphic neuron)。因为酪氨酸羟化酶(tyrosine hydroxylase)的不同而具有性别差异特点。kisspeptin是女性雌激素正反馈调节GnRH的关键因素:
- 用kisspeptin抗血清处理后,雌性大鼠的LH冲动性分泌被完全阻断;
- 雌二醇诱导AVP神经元kiss1mRNA表达;
- AVP神经元kiss1mRNA表达的峰值发生于GnRH/LH分泌冲动时;
- kiss1-雌激素受体阳性神经元与GnRH神经元有突触联系。
如果人类缺乏kisspeptin受体,不会启动青春期发育。在生理情况下,kisspeptin作为青春期发育的守门因子(gate keeper)而发挥作用。增强下丘脑kisspeptin信号可解除青春期发育前的低促性腺激素状态,因而kisspeptin信号增强是个体开始转向青春期发育的最早标志,接着发生脉冲性GnRH释放并启动青春期发育。2003年报道,KISS1R突变引起特发性下丘脑性性腺功能减退症(idiopathic hypothalamic hypogonadism)。
