
基因的概念及性质
基因的概念是随着遗传学、分子生物学、生物化学等学科的发展而不断完善的。基因是生物的遗传物质,是遗传的基本单位,他携带着特定的遗传信息,这种特定的遗传信息由特定的核酸排列顺序所决定,在特定的条件下能表达成特定的性状和生理功能。基因也可以发生突变。
基因具有三个基本特性:
- 稳定性:每个基因都是核苷酸链上的一个特定区段,其在DNA上的数量和碱基顺序上是稳定的,基因的DNA通过半保留复制准确稳定地传给子细胞;
- 特异性:基因通过转录和翻译决定多肽链的氨基酸顺序,从而决定某种酶和蛋白质的性质,最终表达为某一性状,不同的结构基因决定了不同的酶和蛋白质分子,细胞中不同的基因表达导致了细胞分化,决定了细胞的结构特点和功能;
- 可变异性:基因虽然很稳定,但在一定的自然条件下,基因的个别碱基可能被另一种碱基替换,也可能由于一两个单核苷酸的增减而改变整个碱基顺序,从而导致基因功能改变,这些基因分子内的变化总称为基因突变。
基因的结构
基因的化学组成
基因按其功能可以分为结构基因和调节基因两类,前者决定某种多肽链(蛋白质)或酶分子结构,后者指某些可调节控制结构基因表达的基因。此外,还有一些只转录不翻译的基因,如microRNA、piRNA、rRNA和tRNA基因等。
人类的基因主要存在于细胞核内的染色体上,少数存在于细胞质中线粒体的DNA上。细胞核内基因的DNA由编码顺序和非编码顺序两部分构成,编码顺序是不连续的,被非编码顺序隔开。包括人类在内的真核生物基因大小差别很大,平均长度为1~1.5kb,最大的DNA基因全长可达2300kb。
核酸的基本单位是核苷酸,每个核苷酸由1个磷酸、1个五碳糖和1个碱基三部分组成,核酸包括两类:一类是脱氧核糖核酸(DNA),另一类是核糖核酸(RNA)。DNA中的碱基包含四种:即胞嘧啶(C)、胸腺嘧啶(T)、腺嘌呤(A)、鸟嘌呤(G);而RNA中的四种碱基中T被U代替,即RNA包含的四种碱基为胞嘧啶(C)、尿嘧啶(U)、腺嘌呤(A)、鸟嘌呤(G)。
DNA分子是由四种脱氧核苷酸经3′→5′磷酸二酯键聚合而成的多核苷酸链,四种核苷酸的连接和排列顺序构成DNA的一级结构。DNA的空间结构为双螺旋模型,这一结构由Watson和Crick在1953年提出,该学说的要点为:DNA分子是由两条平行的多核苷酸链围绕同一中心轴构成的右手双螺旋结构(B 型DNA)。多核苷酸的方向由核苷酸的磷酸二酯键的走向决定,一条从5′→3′,另一条从3′→5′,两条链呈反向平行排列,彼此由氢键相连,G与C配对,A与T配对。
每个基因的碱基顺序只在一股单链上占有特定的位置,这一股DNA链叫有义链,另一股的同一位置上则是其互补顺序,没有基因的性能,不参与DNA的转录表达。含有特定基因的DNA片段在有关染色体上占有一定的位点,这种相应位点的一对基因称为一对等位基因。在配子的减数分裂过程中,随着同源染色体之间有规律的分离和组合,基因稳定而准确地传递给后代。
DNA双螺旋在一定条件下,互补的两条链的氢键可以发生断裂和重组,因此出现双链和单链的可逆转变。在细胞分裂的过程中,随着染色体的解旋,DNA双螺旋也解旋进行复制,两链各自合成新的互补链,形成新的DNA双螺旋。DNA双链的分合也可在一定的人工条件下进行。DNA水溶液经过高温和碱的处理,便解开成为两条单链,并往往失去以往活性,这个过程叫变性;变性的DNA在缓慢降温的条件下,两条链重新联合,恢复双链结构和活性,这一过程叫复性。DNA双螺旋的变性与复性在分子生物学和分子遗传学研究中应用甚广。根据两条单链复性的快慢,可以计算基因组的大小,因为基因组越大,所含核苷酸越多,复性时间也会越长。依复性时间的快慢也可发现重复顺序的存在,因为多拷贝比单拷贝复性较快。依据DNA的变性复性与互补配对原理,我们可以设计进行核酸分子杂交技术,用以分析检测基因及其突变。
人类结构基因的组成
人类结构基因包括4个区:①编码区,包括外显子与内含子;②前导区,位于编码区上游,相当于mRNA5′末端非编码区;③尾部区,位于mRNA 3′编码区下游,相当于3′末端非编码区(非翻译区);④调控区,包括启动子和增强子等。
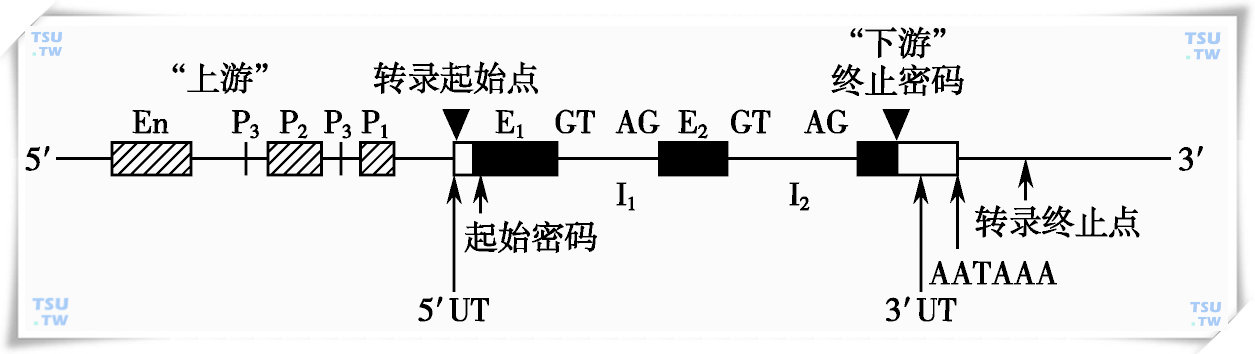
真核生物结构基因的结构示意图
一、外显子和内含子
人类基因与大多数真核生物的基因一样,为不连续基因,也称为断裂基因。基因的编码顺序在DNA上是不连续的,被非编码顺序隔开。编码的顺序称为外显子,是一个基因表达为多肽的部分;非编码顺序称为内含子,亦称为插入顺序。内含子只参与转录,在前体mRNA的加工过程中被剪切掉。
真核生物也有一些基因没有内含子,如组蛋白基因、干扰素基因等,他们大多数以基因簇的形式出现。
二、外显子-内含子接头
真核生物断裂基因的外显子与内含子的接头区都有一段高度保守的顺序,内含子5′端开头的碱基大多是GT,3′端结尾的多是AG,在RNA的加工过程中,内含子的这种特异性结构在细胞核内的sRNA帮助下,被准确地剪切掉。
三、侧翼顺序
在每个基因的5′端和3′端,即在第一个外显子和最后一个外显子的两侧都存在一段不被翻译的非编码区,这两侧的非翻译区称为侧翼顺序。这些区域中含有基因调控顺序,可以调节该基因表达的活性。在这些侧翼顺序中包含有启动子、增强子和终止子。启动子决定基因转录的起始点,转录的起始频率,激活、调节转录的功能;增强子不能启动一个基因的转录,但有增强转录的作用,增强子顺序可与特异性细胞因子结合而促进转录的进行,因此增强子通常具有组织特异性;终止子具有转录终止的功能,其顺序的共同特征是在转录终止点前有一段回文序列,因此终止子转录后形成的RNA具有发夹结构,帮助转录终止。
基因的复制与表达调控
DNA的复制
基因的复制是以DNA复制为基础的。在细胞分裂过程中,DNA准确地进行半保留复制,把遗传信息从亲代传给子代。
因为DNA双螺旋分子的双链是依AT和CG互补的,所以一条链上的顺序可以由另一条链上的核苷酸顺序来决定。DNA复制过程中,首先是氢键的断裂,双螺旋解旋,然后每条链各自以自己为模板,按互补配对的原则,从周围介质中吸收游离的核苷酸,进行氢键结合,在聚合酶的作用下,合成新的互补链,形成新的子代DNA双螺旋。这样新的2个DNA分子与原来的DNA分子的碱基顺序完全一样,每个子代的DNA分子的一条链来自亲本DNA,另一条链则是新合成的,这种复制方式称为DNA的半保留复制,见下图。
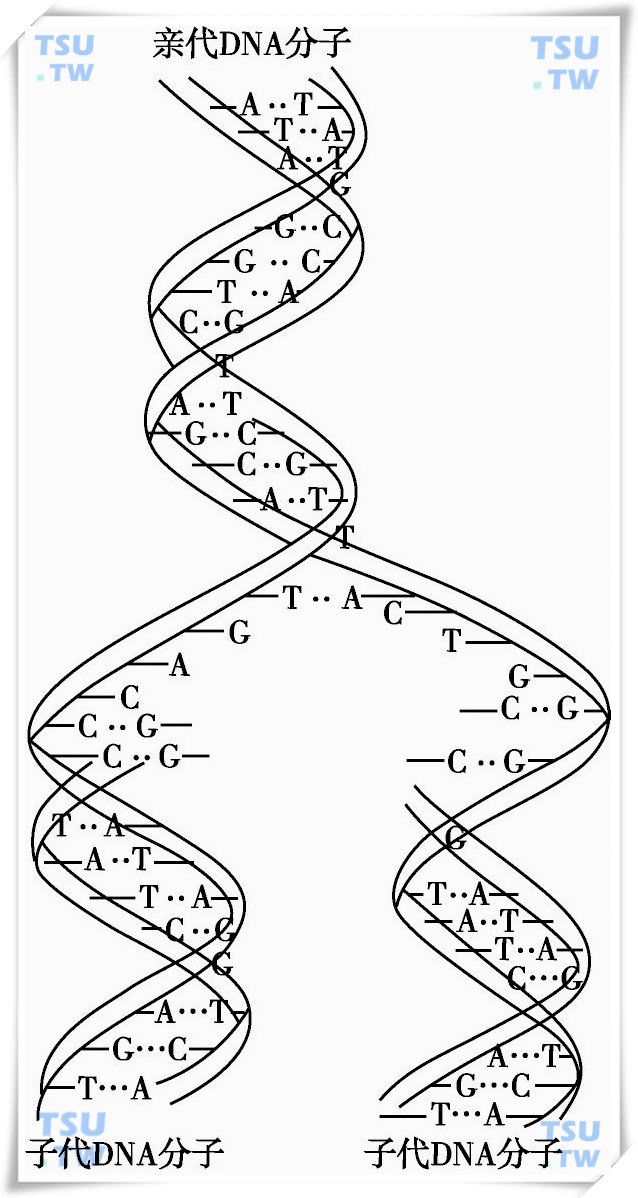
DNA的半保留复制
基因的表达
基因表达是指细胞在生命活动过程中,将储存在DNA中的信息通过转录和翻译,转成具有生物活性的蛋白质分子的过程。
一、转录
在RNA聚合酶的催化下,以DNA为模板合成mRNA的过程称为转录。双链DNA中只有一条作为模板,依A-U和C-G互补的原则转录合成mRNA;另一条互补链不表达转录,但其顺序正好与其对应的mRNA相同,只是DNA中的T变成了RNA中的U,其读码与三联密码对应,因此该不参与转录的这条链也称为编码链。
转录后形成的前体mRNA需经过加工过程,剪切掉内含子,将外显子准确地拼接起来,并经过末端修饰,才能作为蛋白质翻译的模板。
二、翻译
以mRNA为模板,tRNA为运载工具,在有关酶、辅助因子和能量的作用下,将活化的氨基酸在核糖体上装配表达为多肽链的过程,称为翻译。
真核细胞的转录及加工过程都在细胞核内完成,但翻译过程在细胞质中进行。从核糖体上释放出来的多肽链需进一步加工修饰才能形成具有生物活性的蛋白质。该加工过程包括肽链的切断,氨基酸某些基团的羟基化、磷酸化、乙酰化、糖基化等,见图下图。
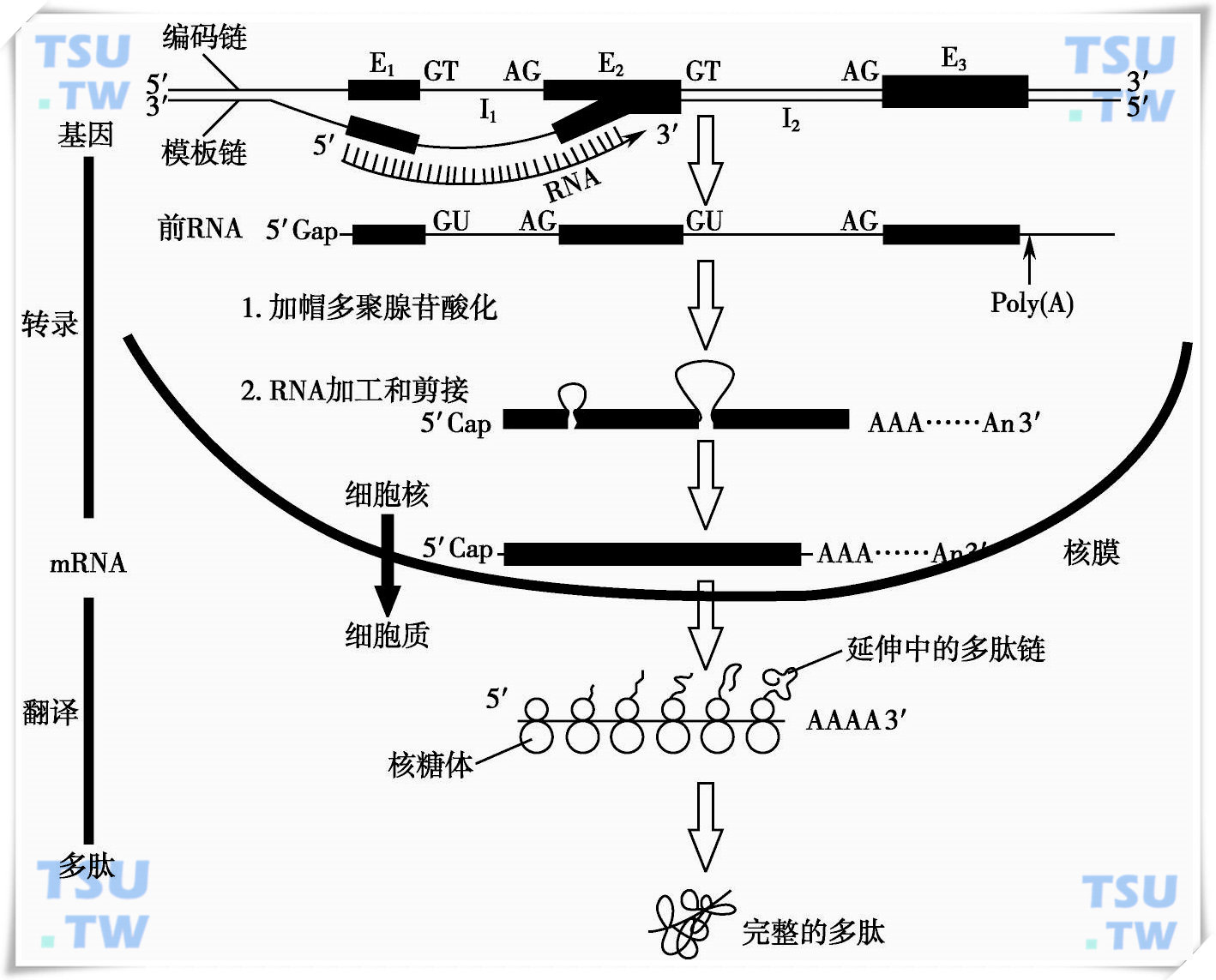
真核生物结构基因表达(DNA→RNA→蛋白质)流程图
基因表达的调控
所有细胞核都是全能的,他们都携带有全部的遗传信息,理论上每一个细胞都有表达的全能性,但实际上并不是全部基因在每个细胞中都表达,也就是说基因的表达具有特异性。特异性包括两个方面:①时间特异性,即基因的表达往往与细胞或个体的特定分化、发育阶段相适应,又称阶段特异性;②空间特异性,即基因的表达是由细胞在各组织器官的分布差异所决定的,故又称为细胞特异性或组织特异性。基因表达的时间和空间特异性在精子发生和成熟的过程中非常明显而且起着非常关键的作用。精子发生就其本质而言,是在严密完整的调节网络(包括内分泌和局部因素等)调控下,特异基因在特定的时空秩序上,精密有序的表达和相互作用的结果。
原核生物的基因表达与调控是通过操纵子机制实现的。1961年Jacob等提出了乳糖操纵子学说,开创了基因表达调控的研究。认为基因的作用单位是操纵子,操纵子的结构是由一个操纵基因和相邻的结构基因构成,他们按一定的线性顺序排列,结构基因的表达产物为特定的酶或蛋白质,操纵基因可以启动全组结构基因的活性,但他又被调节基因激活或抑制。调节基因能合成一种阻遏物,来抑制操纵基因的表达,因此结构基因不能表达;只有当阻遏物被一种特殊的诱导物灭活后,调节基因处于关闭状态,结构基因才能起作用。
相比之下,真核细胞生物的基因调控是多样化调控,更为复杂。包括以下水平的调控,但多数为转录水平的调控。
一、DNA水平的变化
①基因拷贝数的增加、基因丢失、基因扩增,如癌基因的扩增,可导致癌基因的过度表达,使细胞生长分裂失控,导致癌症发生;②基因重排,典型的例子是抗体基因的重排,哺乳动物可以产生108个抗体分子,但人类抗体基因只有约300个,其机制是B淋巴细胞在发育过程中,基因组中抗体基因在DNA水平发生重排,形成编码抗体的完整基因,一个淋巴细胞只有一种组合的抗体基因。因为抗体基因重排中各片段间的随机组合,300个抗体基因足够产生108个抗体。③DNA表观遗传学修饰,如甲基化等,详见相关章节。
二、转录水平调控
主要是蛋白与基因调控序列的作用:①转录因子和启动子是基因转录调控的基本方式,启动子位于基因上游,紧接转录起始点,是转录因子和RNA聚合酶的结合位点;含有TATA盒(RNA聚合酶识别及结合位点)、CAAT盒和GC盒(增强子作用);②转录强化子(增强子)使基因转录只有在适宜转录因子作用下进行,并能对细胞内外的信号作出反应,可以满足细胞分化的需要。增强子常位于启动子上游700bp~1kb处,被转录激活子结合后,改变染色质构型,使DNA歪曲形成环状结构,使强化子与启动子直接接触,便于转录因子、转录激活子和RNA聚合酶一起形成转录复合体,提高转录效率。转录水平的调控还涉及激活因子的种类及调控、选择性启动子、mRNA的切割等方面。
三、翻译水平的调控
①翻译多肽过程的调控,在真核生物的很多生理过程中,转录形成的mRNA被储存起来,在适当的时候才被翻译成蛋白。如海胆卵内的mRNA在受精前不被翻译,在受精后蛋白合成旺盛。其调节的机制是卵细胞内的mRNA仅具有20个核苷酸的多聚A(polyA)尾,只有到了适宜时期,这个尾端序列被加长到几百个的时候,才被翻译成蛋白。在精子发生中,RNA结合蛋白在精子形成期基因的表达也起了重要作用(请参考本章第四节)。②蛋白加工过程的调控,如蛋白的折叠、切割、化学修饰等。
四、microRNA对基因表达的调控
microRNA是近年来发现的普遍存在于生物中的一类非编码小RNA,长度为20nt左右。近年来研究表明,microRNA的主要作用是通过调控基因表达而在生理或病理过程中起重要作用。microRNA主要通过与靶标mRNA互补结合后降解靶标mRNA,达到调控基因表达的作用;microRNA也可通过抑制蛋白翻译而调控基因表达。目前,已在人类发现数百种microRNAs,一种microRNA可以调控多个基因mRNAs的表达;而一个基因的mRNA也可能受多种microRNAs的调控。另外,虽然大部分研究认为microRNA对基因的表达调控是抑制作用,但是有一些研究表明,microRNA对基因的表达调控也可能是促进作用。
以上是表达蛋白的结构基因的表达调控,近年来,对于microRNA、piRNA的表达调控也成为研究热点。目前的结果表明这些小RNA的表达调控与结构基因表达调控有类似之处,有一些microRNA是跟蛋白表达基因一起被转录的。但是,小RNA的表达调控也有其不同的地方,如小RNA稳定性、成熟等水平的调控等。
人类基因组的组成
人类基因组包括细胞核基因组和细胞质中的线粒体基因组。
细胞核基因组
细胞核中的每条染色体包含一个DNA双螺旋分子,因为体细胞为二倍体,所以每个体细胞中有2套染色体(2n),其所含的DNA为两个基因组构成。每个单倍体基因组约含3000Mb,其中编码蛋白质的结构基因只不过5~10万个,仅占基因组的2%~3%,其余的DNA顺序都是不表达的区域,包括基因间的间隔顺序、基因内的插入顺序、重复顺序等。他们的功能目前尚未研究清楚。已定位的基因可以由相关的资料查到,也可在计算机Internet网上获得详尽信息。
线粒体基因组
人类线粒体DNA是独立于细胞核染色体外的一个小基因组,他能半自主地复制,由16 569bp组成,每一个mtDNA分子为环状双链DNA分子,外环为重链,内环为轻链。基因组包含37个基因,其中13个为蛋白质基因,2个为rRNA基因,还有22个tRNA基因。近期,已有学者报道了线粒体中存在microRNA基因。线粒体基因编码的基因主要为与呼吸链相关的一些结构基因,如细胞色素、ATP酶复合体成分、NADH脱氢酶亚单位的基因等。线粒体基因突变导致的疾病多为与神经、肌肉及供能相关的遗传病,自20世纪70年代末开始引起人们的普遍关注。
基因突变
概念
突变是指遗传物质发生的可以遗传给后代的变异。广义的突变包含染色体畸变和基因位点的突变两层意义,两种类型的突变其遗传方式各有特点。通常所说的基因突变主要指狭义的基因突变,包括某一基因的核苷酸顺序或数目发生的改变,仅涉及DNA分子中单个碱基改变者称为点突变,涉及多个碱基突变者包括碱基重复、缺失和插入。这种狭义的基因突变所致的疾病在遗传方式上呈单基因性状遗传,细胞核内染色体上的单基因突变的遗传遵循遗传学三大基本定律;细胞质中的线粒体的基因突变在遗传上呈母系遗传。
体细胞突变和生殖细胞突变的意义
基因突变可以发生在个体发育的任何阶段,可以在细胞增殖周期的任何时期。但发生的时期和部位在遗传学上意义却有不同。如果突变发生在个体形成以后的体细胞中,那么新发生的突变只能传给其子细胞,很少传给后代,通常所见的遗传学效应是在个体的局部产生异常的细胞,导致体细胞遗传病——肿瘤的发生。如果突变发生在生殖细胞如精子和卵子中,则新发生的突变可以通过受精卵传给其子代,若突变基因的性质为显性,则突变的表型在子一代中会马上表现出来;若突变基因的性质为隐性,则会由相应正常显性等位基因的遮盖而不表现该突变基因的性状,子代将成为隐性致病基因的携带者,将来在家系的基因传递中,特别在近亲婚配时,由于隐性致病基因纯合而表现出相应的疾病性状。
自发突变和诱发突变
基因突变既可自发发生,也可因外界因素的诱导产生。因为自然界中诱变因素的作用或由于偶然的复制、转录、修复时的碱基错误配对所产生的突变称为自发突变。人类单基因病绝大部分是自发突变的结果,自发突变的频率很低,平均每世代每核苷酸的突变频率仅为10-10~10-9左右。由于物理、化学因素诱导而产生的突变,称为诱发突变,这些诱导突变的外界因素称为诱变剂,常见的诱变剂有多种辐射射线(如X射线)、重金属离子(如铅和汞等)、某些化学物质(如亚硝酸钠)等。当这些外界因素的接触或使用剂量超过一定标准时,则成为一种诱变剂,可诱发基因突变,导致遗传病或肿瘤发生。
突变热点与频率
从理论上讲,DNA分子的每一个碱基都有可能发生突变,但实际突变部位并不是随机分布的,DNA分子上的各个部位的突变频率各不相同,某些部位的频率远远高于平均值,这些部位称为突变热点。不同基因位点的突变热点构成人群中不同遗传相关疾病发病率的差异。一般而言,生殖细胞的突变频率较体细胞突变频率高,因为生殖细胞在减数分裂时对外界环境具有较高的敏感性。女性卵子发生过程中的基因突变概率较男性精子发生过程中的可能性大,因为卵子的发生过程有一较长的核网期,此时期的染色体呈松散解旋状,易受外界环境中物理、化学因素的影响。
基因突变的种类
按遗传物质DNA的结构改变类型来分,基因突变一般可分为碱基置换突变、移码突变、整码突变及染色体错误配对和不等交换等四种类型。
一、碱基置换突变
遗传物质DNA的组成包含四种碱基:胞嘧啶(C)、胸腺嘧啶(T)、腺嘌呤(A)、鸟嘌呤(G)。一个碱基被另一个碱基取代而造成的突变称为碱基置换突变。其中,同类碱基置换称为转换,即一个嘧啶被另一个嘧啶取代,或者一个嘌呤被另一个嘌呤取代。不同类碱基互相置换称为颠换,即一个嘧啶被另一个嘌呤取代,或者一个嘌呤被另一个嘧啶取代。碱基置换可以在遗传信息的表达过程中导致蛋白质一级结构即氨基酸的组成改变而影响蛋白质或酶的生物学功能。依碱基置换导致氨基酸顺序的影响,又可分为以下几种类型:
- 同义突变:碱基突变后的表达产物中,肽链中的氨基酸没有发生改变,与改变前的氨基酸一样。这是因为遗传密码的兼并性,允许三联密码子的第3个碱基有规律地改变,而且可以编码相同的氨基酸,这种碱基突变并未导致DNA相应遗传信息的改变,即表达产物中的氨基酸没有发生改变,也不会影响相应蛋白质的空间结构,因此将这种碱基突变类型称为同义突变。这种突变不易检出,估计在自然界中这种突变的频率相当高。
- 错义突变:是指DNA分子中的核苷酸置换后改变了mRNA的遗传密码,从而导致合成的多肽链中一个氨基酸被另一个氨基酸所取代,这种碱基突变称为错义突变。在这种情况下,该变异氨基酸的前后氨基酸没有变化,其结果是产生异常蛋白质和酶,也有不少基因由于错义突变而产生部分活性降低和异质组分的酶,因此不完全抑制酶的催化反应。
- 无义突变:在mRNA的表达过程中,有3个密码子的功能是行使终止作用,他们是UAG、UAA、UGA,因为他们没有对应的氨基酸产物,也被称为无义密码。当单个碱基置换导致密码子突变为终止密码时,多肽链的表达将提前终止,所产生的蛋白质或酶大都失去活性或丧失正常功能,这种类型的碱基突变称为无义突变。
- 终止密码突变:当DNA分子中的某一个终止密码发生突变,成为编码氨基酸的密码子时,多肽链的合成将继续进行下去,肽链将延长至下一个终止密码子时才停止,从而形成一条延长的异常的多肽链,这种延长了的多肽链也是功能异常的产物,其功能将部分或全部丧失。这种类型的碱基突变称为终止密码突变。
- 抑制基因突变:当基因内部不同位置上的不同碱基发生了两次突变,其中一次抑制了另一次的突变的遗传效应时,这种碱基的突变称为抑制基因突变。
二、移码突变
指DNA链上插入或丢失1个、2个甚至多个碱基(但不是三联密码及其倍数),在读码时,由于原来密码子移位,导致在插入或丢失碱基部位以后的编码都发生改变,结果翻译出来的氨基酸顺序也发生了相应改变。移码后终止密码会推后或提前出现,造成肽链延长或缩短。
三、整码突变
如 果在DNA链的密码子之间插入或丢失1个或几个密码子,则合成的肽链将增加或减少一个或几个氨基酸,但插入或丢失部位的前后氨基酸顺序不变,这种突变方式称为整码突变,也称为密码子的缺失或嵌入,其插入或丢失的碱基数为3个或3的倍数,为1个或多个三联密码子。
四、染色体的错误配对和不等交换
在配子生成的减数分裂过程中,同源染色体的同源部分会发生联会和交换,如果联会时配对不精确,会发生不等交换,造成一部分基因缺失和部分基因重复。这种突变常用来解释大段核苷酸的丢失或重复。
此外,“动态突变学说”是近年来用以解释一些基因病的理论之一。人类基因组中存在着大量的DNA重复顺序,一些基因编码区或非编码区的CGG、CTG、CAG三核苷酸串联重复序列拷贝数过度增加或扩展,可导致一系列遗传病和肿瘤发生,这种三核苷酸重复片段扩增突变是不稳定的,可以随着世代的传递而扩大,这种三核苷酸拷贝数目的突变称为动态突变。例如脆性X综合征[Fra(X)],即X连锁的智力低下,其发病率:男性1/2000~1/1500;女性1/2500。定位于染色体Xq27.3的FMR1基因突变为该病发生的主要遗传学原因,该基因的5′非翻译区含有三核苷酸重复序列(CGG)n,其动态突变扩展是95%以上的Fra(X)综合征患者发病的分子遗传学基础,前突变n=53~230,全突变n≥230。在前突变状态下,个体无症状或仅有轻微症状。在女性向后代的传递过程中,可累积突变为全突变,这种全突变使其相邻的CpG岛也被甲基化,可关闭相邻基因的表达,从而表现出智力低下等临床症状。
基因突变的后果
根据基因突变对机体的影响程度大小,基因突变的后果可大致分为如下几种情况:
一、不同基因型个体对外界环境的适应能力可不相同,因而导致个体的生存或生育能力的差异。例如镰状细胞贫血症的HbS突变基因的杂合子与正常的HbA纯合子相比,前者更能抵抗疟原虫的感染,不易患恶性疟疾,因此在热带丛林作战时,HbS突变基因的杂合子个体更能适应恶劣的气候,具有更强的生存能力。
二、产生遗传易感性。有些突变基因在个体内的累积,增加了个体易患某种疾病的可能性,即产生了该种疾病的遗传易感性。
三、导致遗传性疾病的产生。突变基因表达出明显的疾病症状,导致个体生存及生育能力降低,寿命缩短。这些疾病包括基因突变所导致的蛋白质异常的分子病和遗传性酶病。
四、产生致死突变,造成死胎、自发流产或出生后死亡。变异后果轻微,对机体不产生明显的效应。从进化的角度来看,这种突变称为中性突变。因为遗传密码的表达中,三联密码的第3个碱基存在兼并性,如果单个碱基的突变正好发生在三联密码的第3个碱基,则该基因的表达产物与正常产物无异,对个体的表型性状不会产生影响。有些错义突变不影响蛋白质或酶的生物活性,因而不会表现出明显的表型性状和活性,这种突变也属于中性突变。
五、造成正常人体生物化学组成的遗传学差异,这种差异一般对人体无影响,仅构成人群的遗传多态性。如人类ABO血型、HLA类型、各种同工酶等。这些个体的差异在一般情况下,对个体的生存及生育无明显影响。但在一些特定情形下,可因为个体的遗传差异而产生严重的后果,如不同的血型间输血,不同HLA配型间的同种移植产生排异反应等。
六、也可能给个体的生存和生育带来一定的好处。
基因分析及诊断
基因分析
长期以来,基因病的诊断主要依靠临床诊断和生化检查,而且检查的主要目标为含有基因表达产物的体液或细胞,受基因表达组织特异性的限制,并要求对基因产物或代谢异常机制有所了解。依目前对遗传病的了解而言,还远未达到这种认识。理想的诊断方法是对患者基因或DNA本身直接进行分析,因为这种分析摆脱了上述各种限制。机体各种组织的有核细胞均含有全能核,有全套基因组DNA,都可以作为分析的材料,而不必考虑表达的问题。如淋巴细胞、皮肤细胞、绒毛细胞、口腔黏膜细胞均可以作为基因分析的便利材料。
基因分析最常用的技术之一是核酸分子杂交技术,其基本原理是:互补的DNA单链能够在一定条件下复性结合成双链,即能进行杂交。这种杂交是特异性的,严格按照碱基互补的原理,不仅能在DNADNA间进行,也能在RNA-DNA间进行。因此,当用一段已知基因的核酸序列作为探针,与变性后的单链基因组DNA接触时,如果两者的碱基完全配对,即互补,从而表明被测基因组DNA中含有已知基因的同源序列。因此,我们知道进行基因分析检测必须具备两个条件:①必须有特异的DNA探针;②必须获取待测的基因组DNA。当两者都经变性呈单链状态时,即能进行分子杂交来检测分析基因。
基因诊断
临床基因诊断常用技术包括如下几种,但随着分子生物学技术的发展,尤其是高通量技术和新技术,以及生物信息学的应用,将会有更多的技术用于临床诊断:
一、Southern印迹杂交
当用某种特定的限制性内切酶消化基因组DNA时,DNA会被切割成大小不等的片段,通过凝胶电泳可以将他们按相对分子质量的大小分开,相对分子质量越小的片段迁移速度越快,反之相对分子质量越大的泳动速度越慢。为了分析的便利,英国科学家Southern首创了印迹法:用对单链DNA分子具有很强吸附力的硝酸纤维滤膜(NC膜)或尼龙滤膜覆盖在电泳后的凝胶上,其上方压上多层吸水纸,借助浓盐液的上吸作用,凝胶上的单链DNA将转移到滤膜上。DNA的转移是原位的,其对应的相对分子质量位置不变。转移结束后,经过高温烘烤的DNA将固定膜上,待检基因便包含于其中。用特定的同位素探针进行杂交,便可将信号显示出来。如果有基因的缺失或突变则可能导致检测带的缺失或位置改变,见下图。
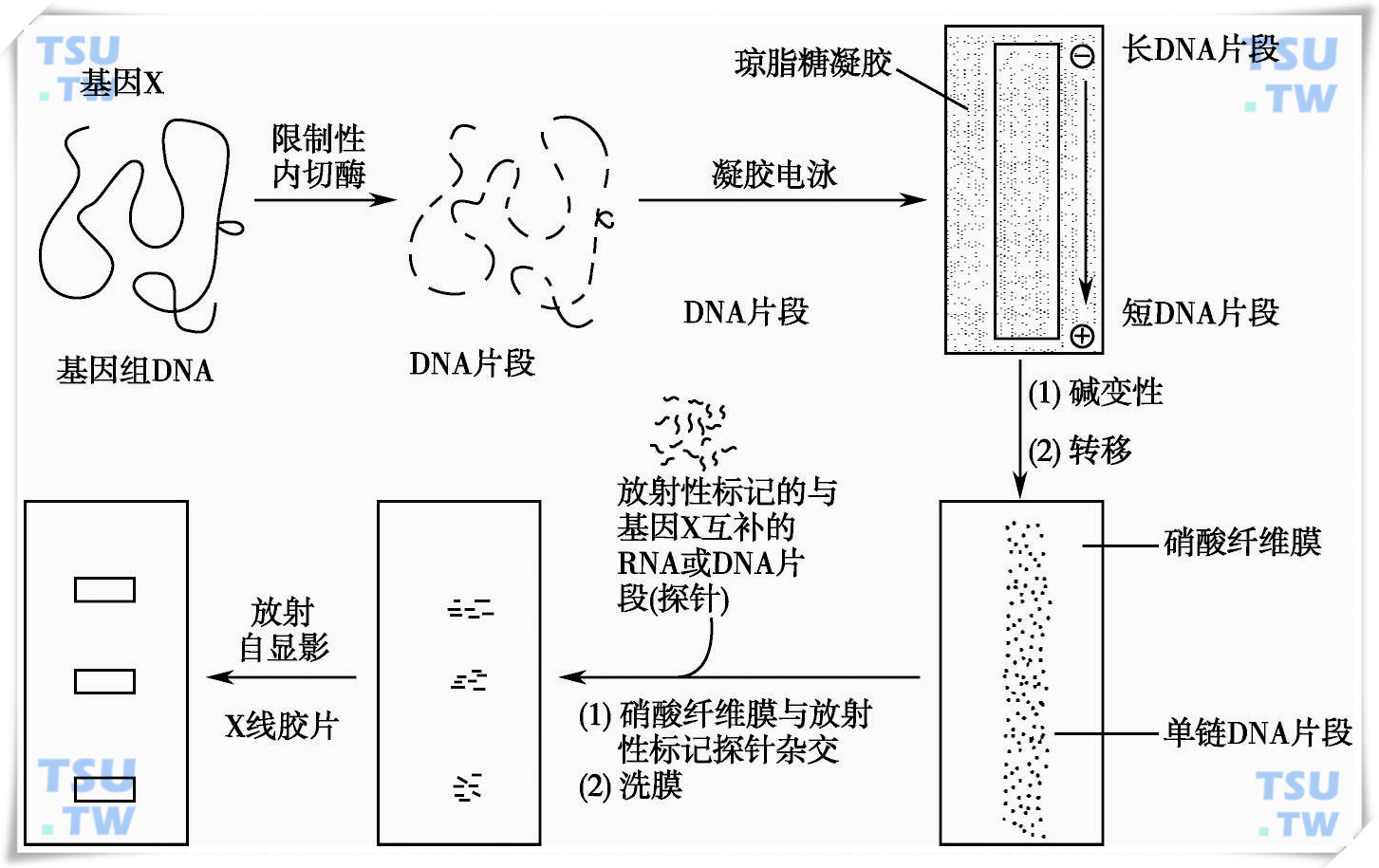
Southern印迹杂交示意图
分子杂交是基因分析诊断的基础,除印迹杂交外,还有斑点杂交法,即将DNA样品变性后直接点在NC膜上,再与探针杂交。
二、聚合酶链反应
近年来,基因分析和基因工程技术有了革命性的突破,这主要归功于聚合酶链反应(polymerase chain reaction,PCR)的发展和应用。应用PCR技术可将待测的目的基因片段在体外扩增数十万甚至上百万倍。这样少量的单拷贝基因通过PCR反应便可直接观察到,原来需1~2周才能作出的基因诊断,可以缩短到几小时便可完成。
PCR反应的基本原理为:在一个体外建立的体系中,包含有一对引物(每个引物的长度为17~20bp左右)、底物dNTP、Taq DNA聚合酶及一定的镁离子浓度,依次设置一定的高温、中温及低温的循环处理,高温使双链DNA变性成为单链,在较低的温度时,引物将与待检DNA链复性结合,在聚合酶的作用下,利用溶液中的核苷酸原料,不断合成新的互补链,这样反复循环25~40个周期,便可以得到大量的DNA片段,见图12-5。
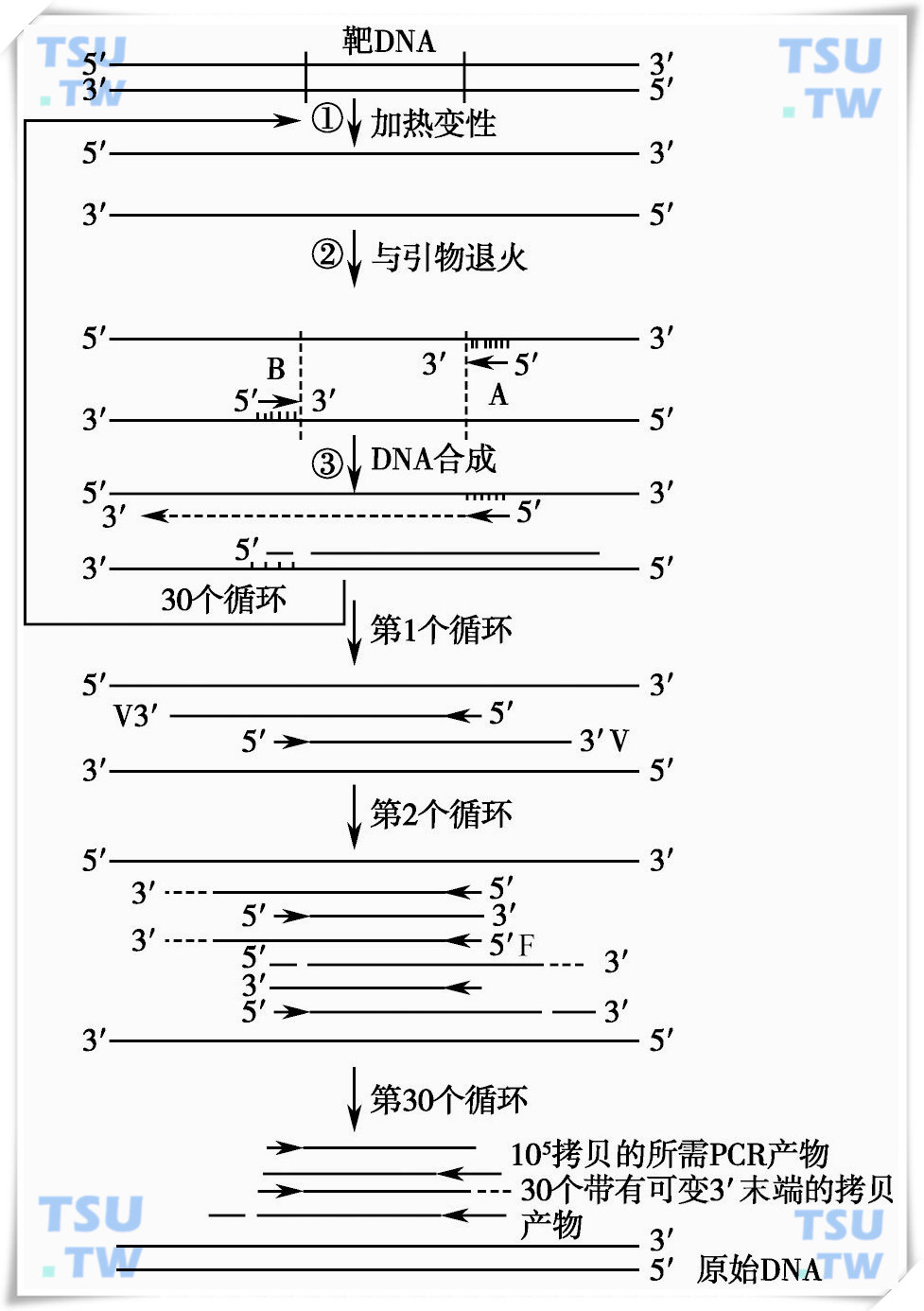
PCR扩增原理示意图
PCR反应特异性强、灵敏度高,极微量的DNA即可作为扩增的模板得到大量的扩增片段,毛发、血痕、肿瘤活检组织、精斑等均可作为鉴定诊断材料。
三、扩增片段长度多态性
小卫星DNA和微卫星DNA的长度多态性可以通过PCR反应后的电泳检测来鉴定,这种方法可用于致病基因的连锁分析,这种方法称为扩增片段长度多态性(amplified fragment length polymorphism,Amp-FLP)连锁分析法。此法多用于突变性质不明的连锁分析。
四、等位基因特异寡核苷酸探针诊断
当基因的突变部位和性质已完全明了时,可以合成等位基因特异的寡核苷酸探针(allele-specific oligonucleotide,ASO)用同位素和非同位素标记来进行诊断。ASO探针一般为长20bp左右的核苷酸,检测点突变时通常需要合成两种探针,一种与正常基因序列一致,能与正常基因稳定杂交,而不与突变基因杂交;另一种探针与突变基因序列一致,能与突变基因杂交,但不与正常基因杂交,这样就可以把只有一个碱基发生了突变的基因区别开来。
五、单链构象多态性
单链构象多态性(single strand conformation polymorphism,SSCP)是指单链DNA由于碱基序列不同可以引起构象差异,这种差异可以造成长度相同或相近的单链DNA的电泳迁移率不同,从而可将发生了单个碱基替代、微小缺失或插入的DNA片段检测出来。
用SSCP方法检测基因的点突变时,通常在疑有突变的DNA片段附近设计合成一对引物,进行待测样品的PCR扩增,然后将产物变性,进行聚丙烯酰胺凝胶电泳,突变所引起的DNA构象差异将表现为电泳带泳动位置的差异。PCR-SSCP方法能快速、灵敏地检测有无突变,但要阐明突变的性质,还需作DNA序列分析。
六、DNA测序
目前,已有多种技术可以用于测定DNA的序列,临床诊断可以根据需要选择实验及测序方案。另外,高通量测序技术的发展,以及成本的大大降低,使得其应用于临床研究,甚至临床诊断,在未来成为可能。
基因治疗
基因治疗是指将外源正常基因导入靶细胞,以纠正或补偿基因缺陷或异常引起的疾病,以达到治疗的目的。通常采用两种策略来达到治疗的目的,其一是纠正突变基因的直接疗法,此为理想的策略,实际上有困难而未实现;其二为用正常基因代替致病基因的间接疗法,此法较为可行,已有一定的临床实践意义。20世纪80年代发展起来的反义技术是在RNA、DNA水平治疗疾病的一种很有前途的高新技术。
依基因转移的受体细胞不同,基因治疗分为两条途径:生殖细胞基因治疗和体细胞基因治疗。
生殖细胞基因治疗
生殖细胞基因治疗是将正常基因转移到患者的生殖细胞(如生精细胞、卵细胞和早期胚胎中),使其发育成正常个体。这是一种理想的方法,但在人类研究中,尚未取得实质性进展,限于其可行性或伦理学要求,目前人类的研究多不考虑该途径。
体细胞基因治疗
将正常基因转移到体细胞,使之表达为基因产物,以达到基因治疗的目的。这种方法的理想措施是将外源正常基因导入靶细胞内染色体上的特定基因座位,用其替代突变基因。该法目前有很大的困难。
目前体细胞基因治疗主要是将基因转移到基因组中的非特定座位,随机整合,只要该基因能有效地表达出产物,便可达到治疗的目的。这种方法不是修复基因结构异常而是补偿异常基因的缺陷,相对而言易于成功。基因治疗中作为受体细胞的体细胞,多采取离体细胞,先在体外导入外源基因,在有效表达后,再输回到体内,这属于间接基因疗法。
体细胞基因治疗不必矫正所有的体细胞,因为每个体细胞都具有相同的染色体,有些基因只在一种类型的细胞中表达,即具有表达的组织特异性,因此治疗只要集中到这类细胞中,如外周血、骨髓等。其次,某些疾病如遗传性酶病,只需少量基因产物即可改善症状,用不着全体体细胞都充分表达。
外源基因的转移有多种方法,常用的有4种。
一、化学法:将正常基因DNA与带电荷物质磷酸钙、DEAE-葡聚糖或与若干类脂质混合,形成沉淀的DNA微细颗粒,使其与受体细胞接触混合,正常基因便有可能转入受体细胞中。该法的缺点是效率低。
二、物理法:包括电穿孔法和直接显微注射法,另外应用人工脂质体包装外源基因,再与靶细胞融合,或直接注入病灶,也是物理法基因治疗的方法之一。
三、同源重组法:是将外源基因定位导入受体细胞的染色体上,通过与同源序列的单一交换或双交换,新基因片段替代有缺陷的基因片段,达到修正基因的目的。
四、病毒介导基因转移:以病毒为载体,将外源基因通过基因重组技术,将其组装于缺陷病毒上,让这种病毒特异感染受体的宿主细胞。目前使用的病毒载体包括有反转录病毒载体和DNA病毒介导载体两种类型。
