
精子发生需要适宜的温度,哺乳动物阴囊温度一般比腹腔内温度低2~8℃,适于精子的生成。几乎所有哺乳动物(除大象,鲸鱼外)的精子发生过程对温度非常敏感。正常的阴囊温度是哺乳动物精子发生所必需的。临床上对少精子症或无精子症的不育男性研究发现,由于内源性的睾丸温度调节失控、如隐睾症、精索静脉曲张或者由于职业原因长期高温作业等,几乎所有患者睾丸温度都异常升高。研究表明,睾丸内温度每升高1℃就会抑制14%的精子生成,精子数量明显减少。若将小鼠的睾丸置于腹腔,精子发生就会停滞,继而将睾丸放回阴囊,精子发生得以恢复。若将大鼠或恒河猴睾丸局部置于43℃水短时处理后,精子发生迅速受到抑制,而经过一段恢复期后,精子发生可恢复到正常水平。
热激诱导生精细胞凋亡
人类的睾丸温度低于体温,用体温或高于体温的温度局部处理睾丸,及隐睾症、精索静脉曲张等均能导致生精细胞死亡增加。对恒河猴和大鼠进行隐睾手术都能引发生精细胞凋亡,并导致无精子或严重少精子。研究结果显示,隐睾手术后,恒河猴生精细胞的凋亡在第7d左右达到高峰,大鼠7.5d左右生精细胞凋亡达到高峰,猴子单侧隐睾手术,术后5d隐睾睾丸中生精细胞发生凋亡,10d后生精细胞大量凋亡,到30d后达到高峰,2个月后精子发生逐步恢复正常,此时几乎检测不到凋亡信号。
隐睾手术对生精细胞的影响
隐睾手术后,随着隐睾天数的增加,隐睾重量呈时间依赖性下降,隐睾第15d时,隐睾重量可降至对照阴囊睾丸的一半。
组织形态学分析显示,隐睾第5d时,生精上皮开始脱落,生精小管开始不完整,但生精上皮的有序排列基本不变。第10d时生精细胞数目明显减少,生精上皮的有序排列被严重破坏,生精上皮可见有多核细胞出现,这是生精细胞发生凋亡的一种早期标志。隐睾第15d时,由于生精细胞不断丢失,生精上皮变薄,生精小管中仅保留Sertoli细胞、精原细胞和少数初级精母细胞,生精小管的直径也明显变小。
TUNEL结果显示,隐睾手术第3d,大量生精细胞凋亡,第7d达到高峰。TUNEL标记为阳性的细胞主要为初级精母细胞和精子细胞。第10d的隐睾随着生精细胞大量丢失,凋亡信号随之减弱。第15d时,由于生精小管中生精细胞已基本上完全丢失,在隐睾中几乎检测不到凋亡信号(下图)。
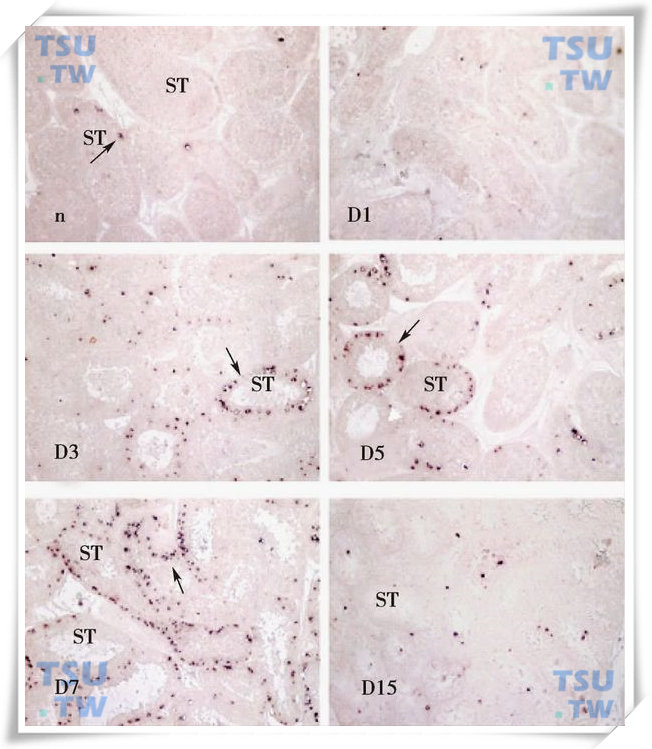
3’末端原位(TUNEL)分析食蟹猴隐睾生精细胞的凋亡
n为阴囊睾丸;D1、D3、D5、D7和D15分别为隐睾后第1到第15d的隐睾。ST:生精小管。箭头所示为凋亡的生精细胞。作为对照的阴囊侧睾丸生精上皮中仅有很少的凋亡信号,隐睾第3d凋亡明显增加,第7d时凋亡信号达到最强,而第15d的隐睾中凋亡信号明显减弱。×200(引自Tao SX et al:Frontier in Bioscience,2006,11:1077-89)。
PCNA是与细胞增殖有关的核蛋白,在核内充当DNA聚合酶的辅基,在体细胞的有丝分裂中,在CDKs及其抑制剂调控下对DNA的复制和修复起着关键的作用,与Ki67同为检测细胞增殖活性的重要标志分子。Ki67和PCNA分析表明,隐睾后期随着生精小管的破坏日趋严重,生精细胞的增殖活性几乎受到完全抑制。说明隐睾导致的生精细胞减少,一方面是由于抑制了生精细胞的分化增殖,另一方面是由于增加了生精细胞凋亡。
43℃水浴局部处理睾丸对生精细胞的影响
43℃水浴局部处理大鼠睾丸,生精细胞凋亡增加,导致生精上皮发生可逆性改变。实验证明用43℃水浴局部温浴睾丸被认为是一种安全可逆性抑制精子发生的方法。通过建立短时43℃水浴局部热处理恒河猴和大鼠睾丸的动物模型,观察睾丸局部温度对精液中精子数量下降和睾丸生精小管精子凋亡的影响,系统研究他的作用途径和分子基础。如图20-10所示,恒河猴睾丸在43℃水浴中局部温浴,每天30min,连续2d,停止温浴。在不同恢复期取精液和睾丸样品,检查精液中精子数和生精小管中精子凋亡数。实验结果与隐睾手术所观察到结果基本相似。43℃水浴局部处理恒河猴睾丸每天30min,连续2d,温浴28d后,精子浓度降为处理前对照组的8.4%,随后精子浓度逐步恢复,到第84d基本恢复到正常水平范围,到第144d后精子浓度完全恢复到正常对照组的水平。
43℃水浴局部处理恒河猴睾丸严重损坏了生精小管的结构和形态,组织形态学分析显示,43℃热处理后第3d生精上皮中生精细胞开始脱落;第8d开始至30d生精上皮呈现退行性变化,表现为生精上皮排列紊乱,出现空泡化,并形成多核巨细胞(生精细胞发生凋亡的一种早期标志),从他的核形态看,可能是来源于多个圆形精子细胞。大多数的生精上皮变薄,管腔内缺失精母细胞和圆形精子细胞,但靠近基底部的精原细胞不受影响;至84d时,大部分生精上皮已经开始恢复,重新出现各级生精细胞有序排列;至144d,生精上皮已完全恢复到处理前的对照水平(下图)。
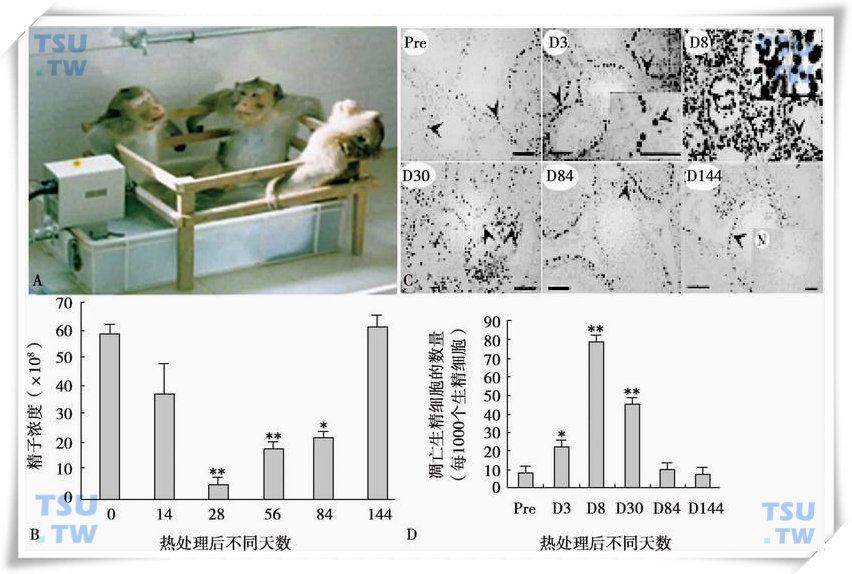
睾丸局部43℃水浴模型中生精凋亡信号
A:猴睾丸局部43℃水浴处理模型;B:43℃热处理后不同天数精子数量的变化;C:3’末端原位分析睾丸局部43℃热处理后生精细胞凋亡信号,pre表示正常睾丸,D3、D8、D30、D84和D144分别为热处理后不同天数的睾丸,N为阴性对照,箭头所示为凋亡的生精细胞,标尺=100μm;D:凋亡信号的统计结果。图中显示的数值为平均数±SEM(n=3),*代表差异显著(P<0.05)或**(P<0.01)(引自Zhang XS et al:Endocrinology,2006,147:1237-1245;Frontiers in Biosciece,2005,10:13110-13121)。
43℃热处理睾丸诱导生精上皮发生凋亡并抑制生精细胞增殖活性,引发生殖细胞的凋亡是周期特异和细胞特异的,在热处理6h内处于生精周期早期(Ⅰ~Ⅳ)和晚期(Ⅻ~ⅩⅣ)的生殖细胞发生凋亡,对热最敏感的生精细胞是Ⅰ-Ⅳ期的粗线期精母细胞和早期精子细胞以及Ⅻ-ⅩⅣ期的粗线期、双线期和分裂期的精母细胞,而Ⅴ~Ⅵ及Ⅶ~Ⅷ期对热不是特别敏感,通过GnRH拮抗剂抑制睾丸内睾酮含量的方法,发现睾丸内的睾酮保护了Ⅶ~Ⅷ期生殖细胞在热处理后的凋亡。
细胞凋亡和增殖在进化上是十分保守的两个过程,他们在正常发育和维持机体的动态平衡中起重要作用。Ki67作为细胞增殖活性的标志分子表达于G1晚期、S、G2和M期细胞的核基质中,在G2期和M期早期表达最强,在G0期细胞和G1早期的细胞中不表达,而且在DNA损伤修复过程中也不表达。免疫组化结果显示,在热处理前睾丸的精原细胞核中检测到Ki67的表达,热处理后被标记的精原细胞几乎检测不到,与隐睾能显著抑制Ki67在精原细胞中的表达一致。这表明热处理后精原细胞的有丝分裂活性明显受到抑制,热引发的少精不仅与细胞凋亡有关,而且精原细胞的增殖活性也受到抑制。
热激对睾丸生精细胞凋亡调控因子的影响
Bcl-2家族是与CED-9同源的哺乳动物蛋白,主要位于线粒体膜上,控制线粒体内细胞色素C的释放,是调控细胞凋亡的重要因子。隐睾能使Bax蛋白从细胞质转向细胞核分布,使Bcl-2蛋白表达升高,与生精细胞的凋亡一致,说明Bcl-2/Bax在隐睾诱导的睾丸生精细胞的凋亡中起着重要作用(下图)。
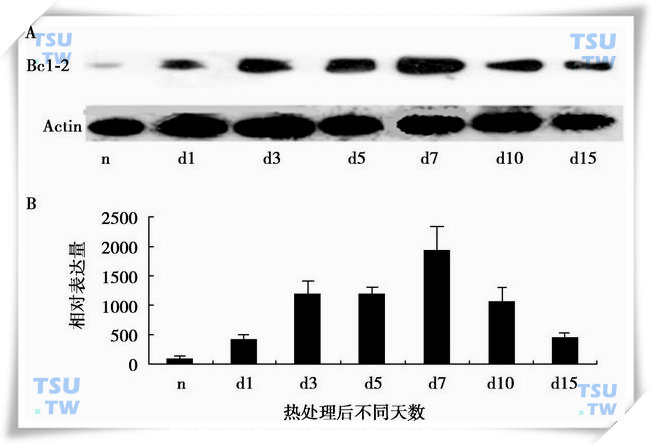
Bcl-2蛋白在隐睾中的表达变化
A:Bcl-2蛋白的Western blot结果;B:Bcl-2蛋白水平的扫描定量分析。其中n为阴囊睾丸;d1、d3、d5、d7、d10和d15分别为隐睾后第1到第15天的隐睾。(引自Tao SX et al:Frontier in Bioscience,2006,Jan 11:1077-89)。
在热处理后30min内生精细胞内Bax从胞质中向核周的重新定位是生精细胞凋亡的诱因。Bax定位的变化引起了细胞色素C的变化从而激活了caspase9以及caspase3、6、7。在Fas-L缺失小鼠中没有发现热引起生精细胞凋亡的阻碍,说明热诱导的生殖细胞凋亡主要是通过线粒体依赖的以及可能的内质网依赖的通路实现的。
在隐睾术后5d,生精细胞发生凋亡的早期,HSP70-2表达几乎不变,但术后10d伴随生精细胞的凋亡丢失,HSP70-2表达显著降低。上述结果提示,在实验性隐睾的早期阶段,HSP70-2可能没有参与抑制生精细胞凋亡过程,他不属于热激诱导的生精细胞早期凋亡相关基因。另外,热激后睾丸中HSP60蛋白表达明显升高是与Bcl-2表达增加有关。因此推测,隐睾生精上皮细胞中HSP60表达的增加可能是通过与Bax形成复合物,将Bcl-2释放出来,由此通过线粒体途径促进生精细胞凋亡。
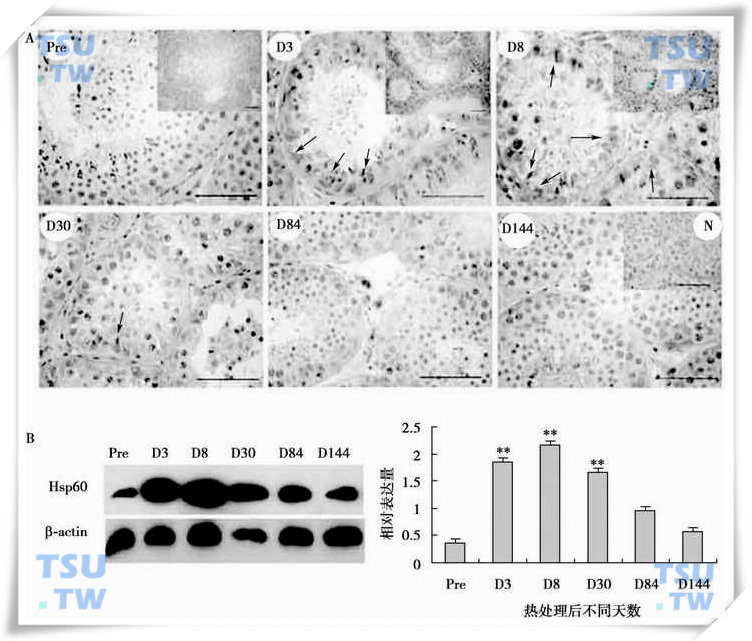
HSP60蛋白在热处理猴睾丸中的表达
A:HSP60蛋白在猴睾丸中的免疫组织化学定位,Pre表示正常睾丸,D3、D8、D30、D84和D144分别为热处理后不同天数的睾丸,N为阴性对照,标尺=100μm;B:HSP60蛋白的Western blot结果及相对表达量的统计分析。图中显示的数值为平均数±SEM(n=3),**代表差异显著(P<0.01)。(引自Zhang XS et al:Frontiers in Biosciece,2005,10:3110-3121)。
HSP60主要定位在精原细胞和Sertoli细胞中,热处理能上调他的表达,提示HSP60可能也起到了保护精原细胞免于热损伤的作用(上图)。HSP105主要在热处理前食蟹猴生精上皮的精子细胞中表达,热处理下调了他的表达水平,但热处理后第3、8d发现有部分精原细胞也表达HSP105,提示HSP105可能通过诱导精子细胞凋亡,而保护精原细胞免于热损伤(下图)。
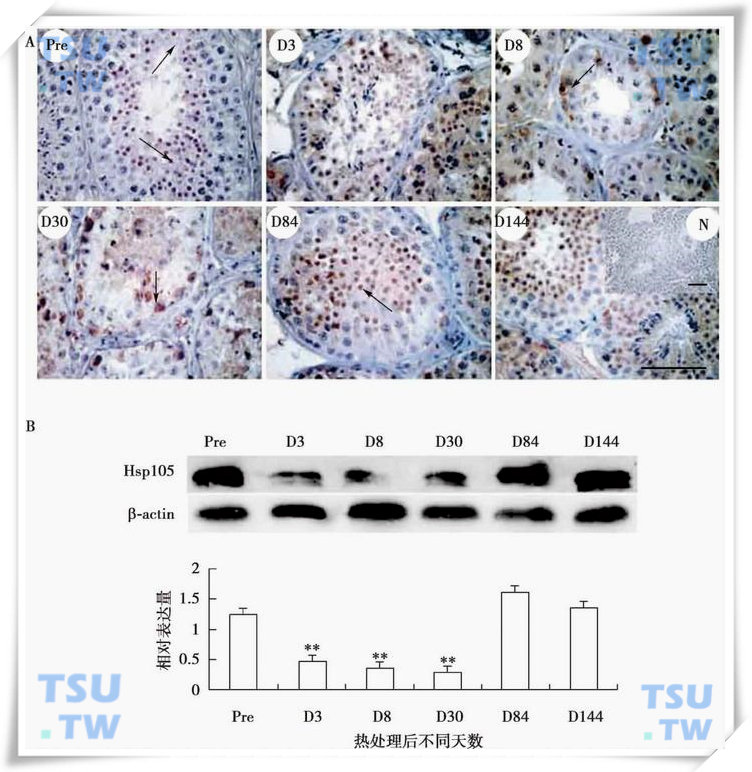
HSP105蛋白在热处理猴睾丸中的表达
A:HSP105蛋白在猴睾丸中的免疫组织化学定位,Pre表示正常睾丸,D3、D8、D30、D84和D144分别为热处理后不同天数的睾丸,N为阴性对照,标尺=100μm;B:HSP60蛋白的Western blot结果及相对表达量的统计分析。图中显示的数值为平均数±SEM(n=3),**代表差异显著(P<0.01)。(引自Zhang XS et al.Frontiers in Biosciece,2005,10:3110-3121)。
另有文献报道,睾丸温度的升高可以诱发p53与HSP105形成复合体,从而稳定p53于胞质中,避免p53入核发挥凋亡作用。因为非活化的p53是存在于胞质中,远离核作用位点。从这个意义上讲,HSP105起到了稳定p53的作用,从而在热处理中可能发挥抑制精子细胞凋亡的作用。43℃局部处理恒河猴睾丸不同时间p53 mRNA表达模式(下图)。小鼠睾丸受热后活化HSF1,但生精细胞中HSP27、HSP70.1以及HSP110并不被诱导合成。HSF1基因敲除小鼠,睾丸热激后粗线期精母细胞凋亡明显受到抑制,而正常对照小鼠粗线期精母细胞凋亡增加。说明HSF1促进热激处理后粗线期精母细胞的凋亡。而与之形成鲜明对比的是,他对精原细胞受热后具有保护作用。这说明HSF1在生精细胞中具有双重作用,这取决于细胞内生存信号和死亡信号之间的平衡。
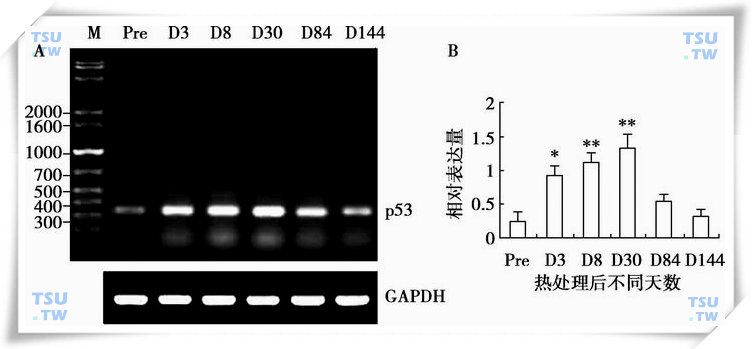
p53mRNA在热处理睾丸中的表达
A:p53mRNA在热处理睾丸中的RT-PCR结果,GAPDH为内参,Pre表示正常睾丸,D3、D8、D30、D84和D144分别为热处理后不同天数的睾丸,M为1kb plus DNA marker(bp);B:p53mRNA相对表达量的统计分析HSP60蛋白的Western blot结果及相对表达量的统计分析。图中显示的数值为平均数±SEM(n=3),*代表差异显著(P<0.05)或**(P<0.01)。(引自Zhang XS et al:J Androl,2006,27(3):405-13).
NO是一个气体自由基分子,由一氧化氮合成酶(NOS)催化合成。NOS家族包括三个成员:诱导型(iNOS)、内皮型NOS(eNOS)以及神经元型NOS(nNOS)。NO和精子发生有着密切的关系。Zini等的研究发现,大鼠经720度扭转精索造成缺血3h后或手术诱导隐睾导致睾丸内生精细胞凋亡时,发现凋亡的生精细胞中有内皮型一氧化氮合成酶(eNOS)的高度表达。研究发现,热激能明显上调诱导型一氧化氮合成酶(iNOS)的表达,而对内皮型一氧化氮合成酶(eNOS)没有明显影响,睾酮也有类似作用(下图)。提示热浴处理和睾酮导致生精细胞凋亡iNOS高表达,表明iNOS可能通过破坏血-睾屏障和黏附连接,参与其中的调控过程。
孤儿受体TR2mRNA主要定位在精母细胞中,热处理并没有显著改变他的表达模式;而他的同源家族成员TR4 mRNA在热处理前的睾丸定位在精母细胞和精子细胞中,热处理使其表达下调。p53 mRNA则在受热处理后的睾丸精母细胞中强烈表达,结合已有文献报道,提示p53可能参与了TR4介导的生精细胞凋亡调控。TR3 mRNA则定位在精母细胞和精子细胞中,热处理能够下调他在这些细胞中的表达。而这一表达模式类似于核转录因子E2F1,已知核转录因子E2F1启动子区有TR3的应答元件结合位点,提示热激通过下调TR3的表达进而抑制E2F1的表达,从而抑制了E2F1的凋亡诱导作用。
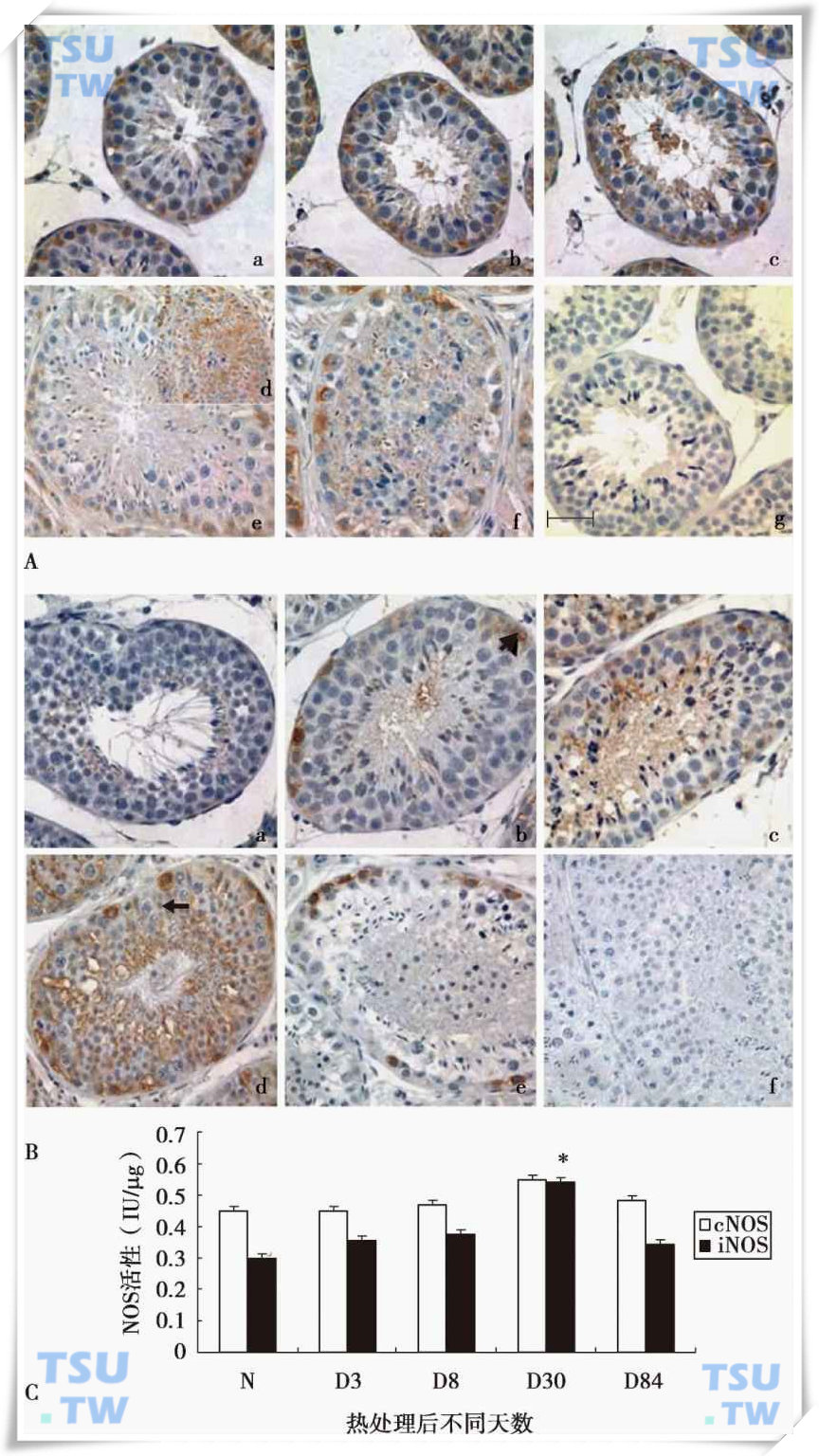
43℃水浴热处理后食蟹猴睾丸NOS的变化
A:水浴热处理后eNOS在睾丸的定位;B:水浴热处理后iNOS在睾丸的定位,a为正常睾丸,b、c、d、e为43℃水浴热处理后第3、8、30和84天,f为阴性对照。
和
分别代表精原细胞和Sertoli细胞。标尺=50μm。C:NOS活性检测。固有型(cNOS)和诱导型(iNOS)。图中显示的数值为平均数±SEM(n=3),*代表差异显著(P<0.05)或**(P<0.01)。(引自Guo J et al.J.Androl,2009,30(2):190-9)
LRH-1称为肝受体类似物1,是与SF-1关系最近的Ftz-F1超家族的孤儿核受体。有证据表明LRH-1在大鼠睾丸的生精细胞和Leydig细胞表达,Sertoli细胞没有表达。实验性隐睾能上调LRH-1在食蟹猴睾丸中的表达。在食蟹猴正常睾丸中,LRH-1在圆形精子细胞和Leydig细胞表达,Sertoli细胞没有表达,43℃局部温浴睾丸后,LRH-1在Sertoli细胞中开始表达。随着生精上皮中生精细胞凋亡,LRH-1表达明显上调,随后生精上皮开始恢复,LRH-1表达下调,恢复正常水平(下图),提示LRH-1可能和生精细胞凋亡有关。
近期有学者报道,43℃热处理睾丸30min后会增高缺氧诱导因子Hif1amRNA的表达,其蛋白转移到生殖细胞核中。同时由于氧化压的增高引起了亚铁血红素加氧酶-1、谷胱甘肽过氧化物酶和谷胱甘肽转移酶表达的增强,温度的升高可能引起了一些复杂的应激反应,包括氧化压和缺氧反应,从而也诱发了生精细胞的凋亡。
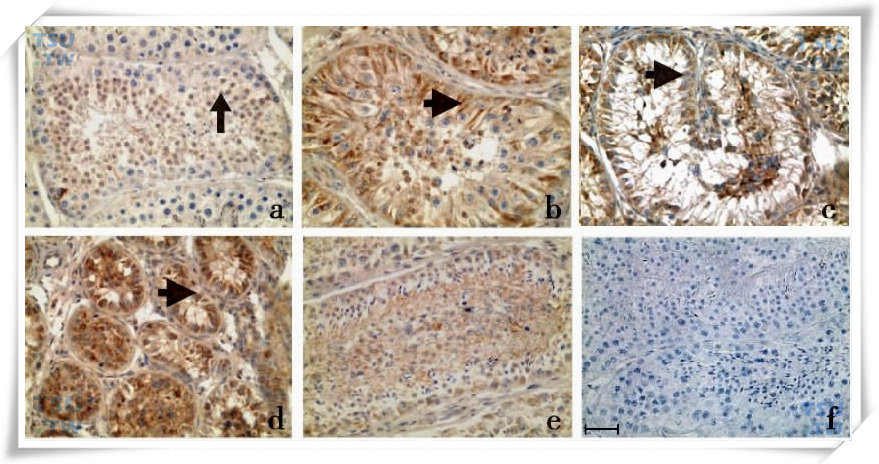
食蟹猴睾丸局部43℃水浴后LRH-1的变化
a为正常睾丸,b、c、d、e为43℃水浴热处理后第3、8、30 和84d,f为阴性对照。 和
和 分别为Sertoli细胞和圆形精子细胞。标尺=50μm。(引自Guo J.et al:. Endocrinology,2007,148(3):1255-65)
分别为Sertoli细胞和圆形精子细胞。标尺=50μm。(引自Guo J.et al:. Endocrinology,2007,148(3):1255-65)
热激睾丸引发基因组表达改变
隐睾导致生精细胞凋亡的上游因素
一、隐睾后基因表达的变化
通过寡核苷酸芯片对隐睾的研究显示(下图),隐睾手术后前4d,明显变化的基因以上调为主,到隐睾术后7d,这种上调趋势改变,代之出现的是大量基因剧烈下调。这一发现是同前人的报导相符的。然而,在隐睾后14d又迎来了一个基因上调峰,到隐睾28d时基因表达出现了最大的下调峰,这表明此时的隐睾在生理上已发生了严重损坏。通过寡核苷酸芯片对隐睾手术后分析显示,热激对精子发生的影响可能是一种强烈的氧化还原代谢应激反应;热激引发睾丸生精细胞大量凋亡是机体试图通过减少睾丸的生精数量以加强热激引起的睾丸对抗氧化还原能力的代偿性需求。我们的实验证实,隐睾手术后首先引发氧化还原相关分子的变化和线粒体相关基因表达的改变,继而引发糖类代谢加强,随后增加脂类代谢加强,以适应新环境的改变,最终导致生精细胞大量凋亡。
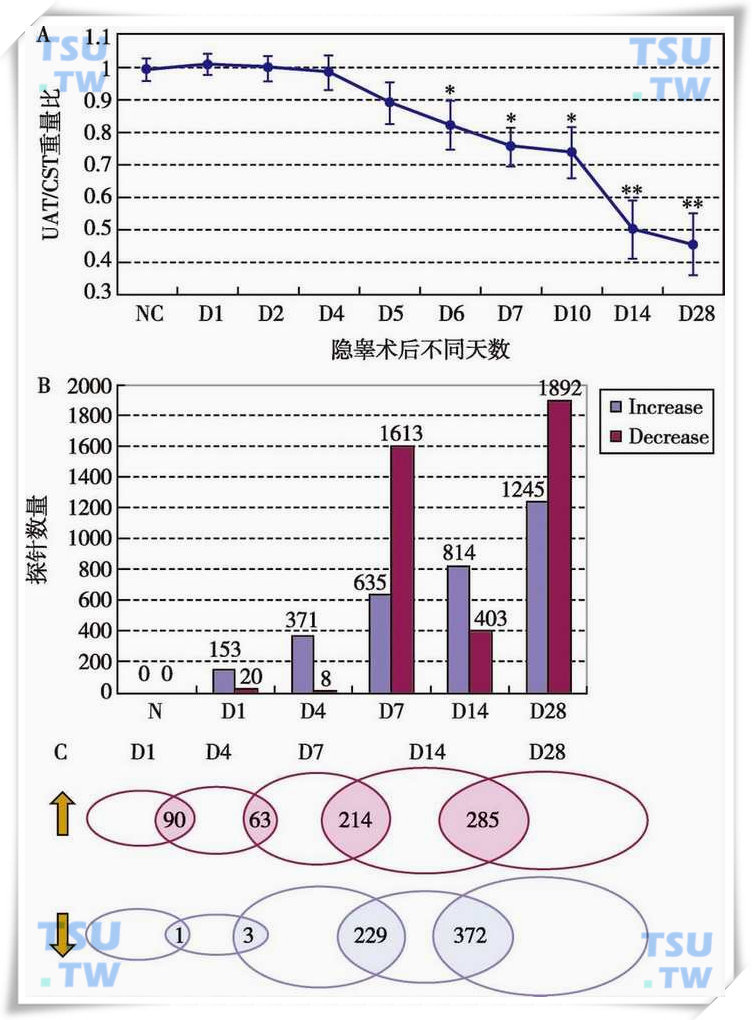
小鼠隐睾睾丸重量及其基因表达模式的变化
A:实验性隐睾(EC)手术后1,2,4,5,6,7,10,14,28d的CD-1小鼠单侧腹腔内睾丸(UAT)与对侧阴囊内睾丸(CST)的重量比。NC为正常对照组。显示的数值为
±SD。*表示与正常对照组相比差异显著(P<0.05);**表示(P<0.01);B:EC后1,4,7,14,28d,基因表达显著上调(荧光信号强度比值的对数(SLR)≥1)和下降(SLR≤1)的基因数量。Increase表示表达上调,Decrease表示表达下降;C:图中重叠圈内的数值表示相邻处理组别的基因表达显著改变的探针数目。每个圈的大小代表各个处理组探针的数目。向上箭头表示表达上调的探针,向下箭头表示表达下降的探针。(引自Li YC et al:Frontiers in Bioscience,2006,11:2465-2482)
二、隐睾手术后最初阶段氧化还原代谢相关基因表达增强
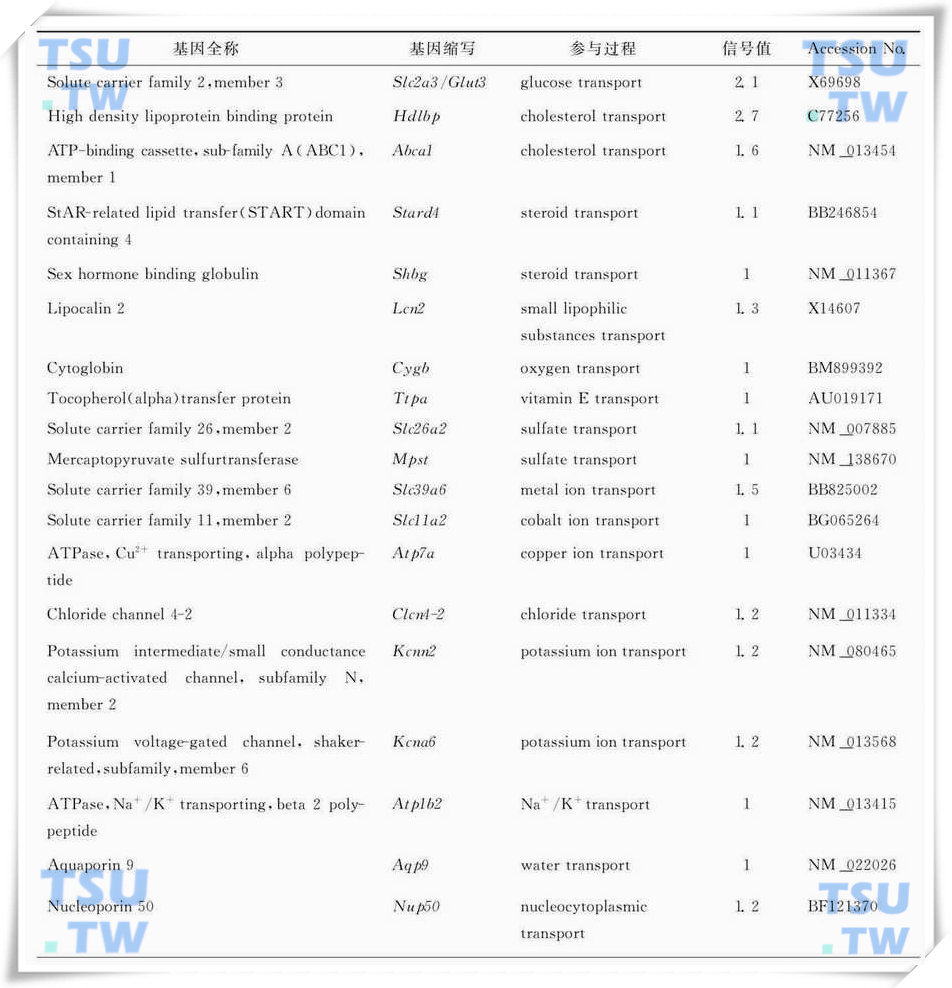
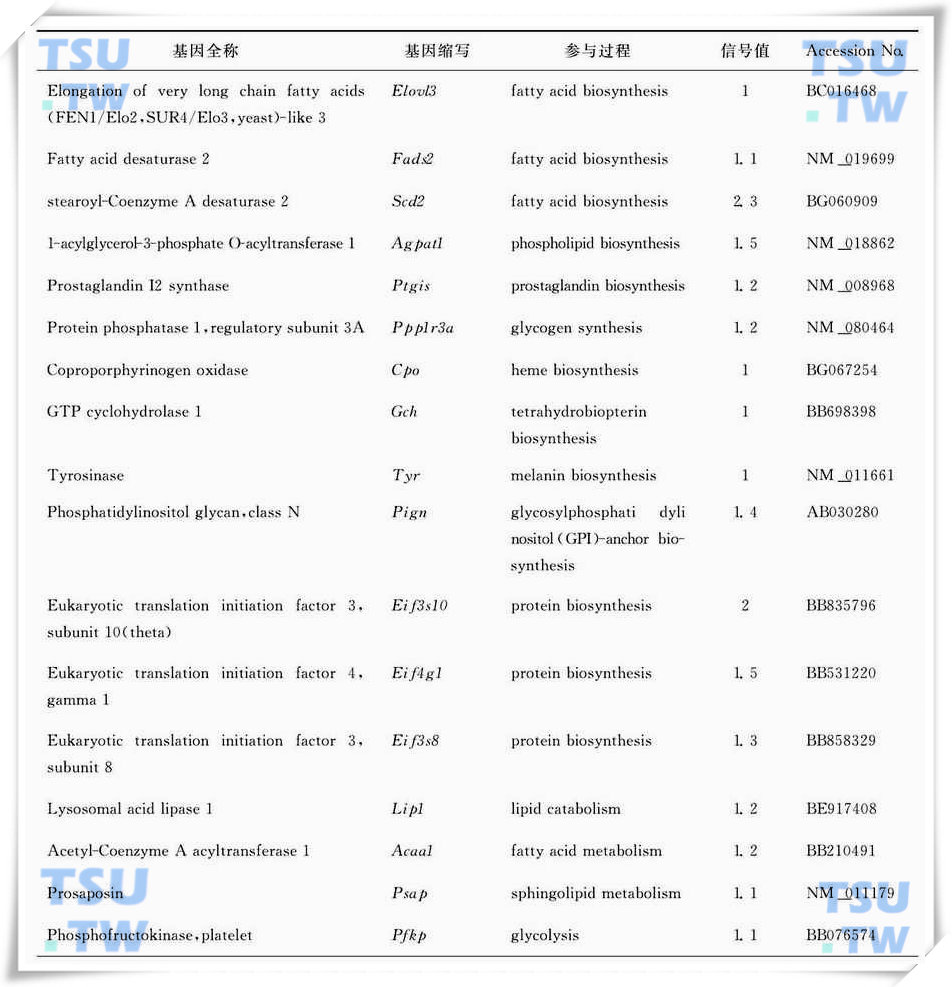
隐睾手术后氧化还原生理代谢活动增强,包括氧、糖、脂、无机盐、水等代谢底物的转运相关基因的转录活性增强(表1),其中最具有代表性的就是葡萄糖转运载体(Slc2a3/Glut3)和高密度脂蛋白结合蛋白(Hdlbp);另外还包括代谢相关酶类的转录活性增强(表2)。
表1:隐睾手术后4d明显上调代谢底物运输相关的基因列表(引自Frontiers in Bioscience 2006,11:2465-82)
 :
:表2:隐睾手术后4d明显上调代谢相关酶类的基因列表(引自Frontiers in Bioscience 2006,11:2465-82)
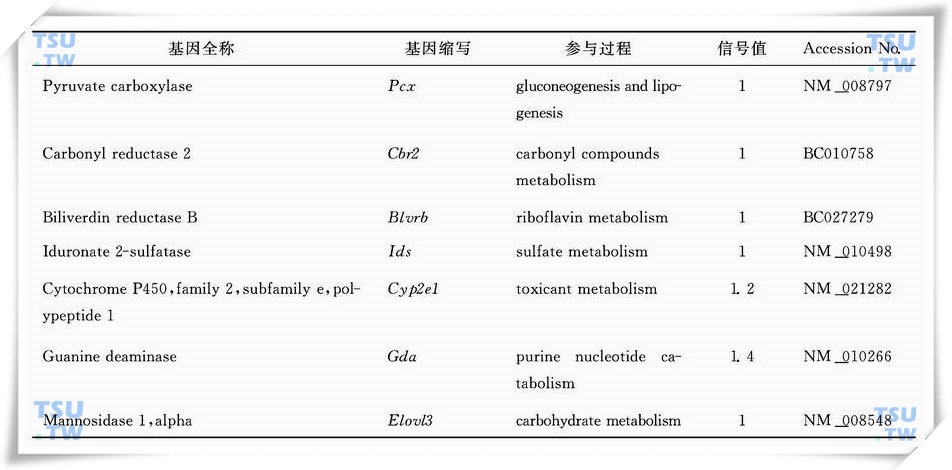
三、隐睾导致氧化应激,代谢改变,抗氧化能力不足
氧化应激增强与其相关基因表达的波动在时间上高度一致。在术后第4d隐睾显示出更有效的抗氧化应激能力,从第5d开始,抗氧化应激能力突然降低,到10~14d又逐渐恢复,28d再次降低。在隐睾手术后第4d,隐睾中多基因高表达,在第5d氧化应激水平上调与第7d基因表达突然下降密切相关。芯片数据表明,高水平的活性氧不仅与其直接来源的异常及其功能紊乱相关,而且与能量代谢的上游生理活动,如脂质代谢和细胞周期调控的异常相关。
隐睾前4d的代谢活动升高体现在糖、脂肪类等能量代谢底物和离子转运的增强,相关的酶类也上调表达,这是代谢增强的一个重要标志;随着隐睾中生精细胞大规模凋亡,糖类转运和代谢相关基因表达开始选择性下调,其中最明显就是生精小管内细胞的主要葡萄糖供应载体Slc2a3/Glut3。已有报道证实,他与生精细胞凋亡有关。他最初明显上调,预示着生精细胞和Sertoli细胞对葡萄糖的大量需求,但隐睾7d 后Slc2a3/Glut3表达开始明显下调;类似的基因还有乳酸转运载体Slc16a7/MCT2,限速酶HK1等。这些基因的下调可能是由于生精细胞的凋亡引起的。隐睾手术后最初阶段代谢活动的增强和睾丸不能有效的控制代谢水平的突然增加,可能引发较高的自由基产生;隐睾发生第一波生精细胞凋亡后(隐睾术后7d),可能糖类供给和代谢不能满足生精细胞的需要,因而增加脂类合成代谢,这可能又进一步加重了抗氧化剂(如NADPH)的生成和分配的不足,同时在隐睾10、14d后,基因的表达上调没有得到合理控制,这可能是导致隐睾7~28d抗氧化能力无法恢复到正常状态的原因。综上所述,隐睾导致生精细胞凋亡的上游因素可能是代谢失控。
精子发生特异相关的新基因
精子发生是一个复杂过程,涉及生精细胞的分化,程序性表达各种细胞特异或阶段特异的基因,在这个过程中,圆形精子细胞调控表达大量与转录、形态结构改变相关的基因,最终形成的成熟精子。圆形精子细胞的形态改变涉及精子顶体、精子核、精子尾、精子细胞质和细胞器等的形态结构变化。从精母细胞的减数分裂、基因重组到单倍体生精细胞染色体重构与浓缩、顶体形成、精子尾的发生等都需要一个精确而有序的调控机制。对精子形成过程机制的解读需要破译大量基因的功能,而这个独特过程又涉及大量新基因。我们利用隐睾模型筛选和克隆了几个与精子发生相关的特异新功能基因:顶体形成相关因子(acrosome formation associated factor,Afaf),睾丸特异性钠氢交换蛋白(testis-specific sodium-hydrogen exchanger,tsNHE),温度相关序列4(temperature related sequence 4,Trs4)和T6441。
一、Afaf
Afaf在圆形精子细胞中大量表达,并且在顶体形成过程中定位于精子细胞顶体内膜和外膜上,可能是膜泡系统的一个新成员。Afaf随细胞周期和状态不同,在质与核中”穿梭”,可能参与细胞的吞噬。Afaf两个不同转录本与GFP融合转染NIH 3T3细胞,7~14d大约有60%的阳性细胞的质膜体积变大,呈显巨大扁平状贴附生长;过表达导致细胞内的膜泡在核周或高尔基体、内质网间隙中不同分布,可能引起膜泡系统转运的紊乱;将Afaf转染HeLa细胞后,其蛋白主要定位于细胞膜和EEA1呈阳性的早期内吞体中,偶尔也可在细胞核中观察到。因此推测,早期内吞体和质膜可能参与了顶体发生。该基因可能是一个与细胞内膜泡形成或转运相关的重要基因,是控制细胞特殊膜泡状态的开关基因。利用链球菌溶血素O型通透性和体外受精实验,进一步探讨了Afaf抗体对受精和顶体胞吐过程的影响。Afaf参与了钙触发的顶体胞吐过程,用Afaf抗体处理过的精子能显著抑制精子对卵的穿透力,从而降低体外受精率。可见,Afaf在顶体胞吐过程和受精过程中发挥重要作用。
二、睾丸特异性钠氢交换蛋白(tsNHE)
钠氢交换蛋白(NHE)家族对于胞内pH值调节具有重要作用,其家族成员在穿膜区具有较强的保守性,他可能是精子活力和获能的一个关键调节子。小鼠tsNHE基因全长1698bp,编码566个氨基酸,理论相对分子质量为62 100,等电点为6.89。利用多种生物信息学软件pfam、SMART等分析,均发现tsNHE基因编码的蛋白质存在钠氢交换蛋白跨膜区的结构功能域。tsNHE蛋白在成年小鼠睾丸的生精小管中特异分布于精子细胞和延长型精子,在成熟精子中特异表达于精子尾部主段,预测其包含12个跨膜结构域且C端无胞内段。针对跨膜区制备的多克隆抗体处理精子,显著降低了精子活力、顶体反应和体外受精率。向精液中加入该抗体,细胞内pH和钙离子浓度均下降。为了进一步探讨tsNHE抗体对生育率的影响,将tsNHE基因疫苗通过肌肉注射、口服或滴鼻法免疫雌性动物,免疫后雌鼠产仔数和生育率与对照组相比显著降低。其cDNA疫苗免疫的雌鼠血清或阴道液可特异性识别精子主段并引发精子凝集。结果显示,其tsNHE抗体能显著抑制体外精子活力和受精。tsNHE可能为开发一种新的男性避孕方法提供一个很好的靶点。
三、温度相关序列4(temperature related sequence 4,Trs4)
Trs4基因全长3110bp,编码框是2373bp,编码790个氨基酸,对温度变化高度敏感。TRS4蛋白具有高度保守性,从果蝇等低等无脊椎动物到人,都有其同源基因。该蛋白含有一个钙调素结合结构域IQ结构域,其C端内含一个WSPW motif,推测这个区域可能是接头部分或功能活性区,推测TRS4的主要功能是参与对钙-钙调素的调控或者介导钙-钙调素的信号。我们克隆并表达了该基因的部分序列,所获得的蛋白免疫家兔获得了该基因的多克隆抗体,免疫组织化学证明TRS4是定位在晚粗线期精母细胞到Step 10精子细胞的核内。稳定转染过表达该基因对Hela细胞没有毒性,在细胞中的分布与细胞骨架相关。我们的结果提示,TRS4可能是一个新的钙调素结合蛋白(CaMBP)对生精细胞的钙离子平衡和基因的转录起着重要的作用。通过多组织RT-PCR、Northern blot、原位杂交证明,Trs4是一个生精细胞特异基因,在晚期粗线期精母细胞、圆形精子细胞、刚开始核变形的精子细胞的核内表达,可能参与减数分裂末期和精子变态之前的生精细胞分化和基因的转录调控,此基因能够对热激作出迅速反应。利用酵母双杂交系统,鉴定出在小鼠睾丸内,TRS4可与Ddc8、Rshl-2、Gstmu1等基因产物相互结合。进一步研究表明,Ddc8是一个功能未知的新基因,在小鼠睾丸中与Trs4的表达类似。Trs4、Ddc8和Rshl-2共同定位于成熟精子的尾部。这些结果表明,TRS4可能通过与多种结合蛋白相互作用,在精子发生的不同阶段,参与精子发生和受精过程。
四、T6441
T6441基因是我们克隆和命名的新基因,全长664bp,编码149个氨基酸,其N端的90个氨基酸十分保守,在大鼠、小鼠、人、猩猩、牛中都有相似序列。大鼠睾丸的T6441基因有4个转录本,4种产物的表达时空相似,可能参与同一种功能和同一个生物过程。其mRNA特异定位于精子细胞,蛋白特异性的定位在10~19期的精子细胞质中,12~19期表达最高,可能参与精子变态的调控。在19期的精子细胞中,T6441的蛋白主要定位在残余小体中,瞬时转染过表达T6441与绿色荧光蛋白的融合基因导致Hela细胞在48h内死亡,提示该基因可能参与了精子细胞质的移动和精子细胞的变态过程,可能与细胞质的脱落有关,其具体的功能尚需进一步探索。T6441基因具有多种翻译产物、多种组织表达的特点;睾丸表达最高,有4种睾丸特异的翻译产物。
上述的4个基因Afaf,tsNHE,Trs4和T6441都在精子细胞中特异表达,且与精子的功能,如顶体反应、获能、精子活力和受精密切相关。局部干扰这些基因的表达不会影响人体健康,因此他们有望成为研发安全有效的男性避孕药的靶点。
综上所述,温度的异常变化会引起生精细胞的凋亡,破坏精子发生过程。同时,温度升高或降低还能影响睾丸中其他细胞的形态和功能。隐睾患者表现为低水平的inhibin B和高水平的FSH,实验证明,双侧隐睾比单侧隐睾具有更高的FSH和LH水平。Kerr等人发现隐睾手术后会使间质细胞形态和功能发生改变,隐睾后间质细胞体积增大,LH受体表达降低。此外,热激后支持细胞的形态和功能都发生了改变。在精子发生过程中,支持细胞对生精细胞的发育成熟提供物理支撑、特殊环境、营养供应、细胞因子分泌和形成血-睾屏障等方面的影响。高温引起的支持细胞形态和功能的改变也会影响精子发生的正常进行。
