
长期以来,人们一直关注温度引起生精细胞凋亡的分子机制。事实上,高温虽然不会诱导支持细胞凋亡,但明显影响他的结构和功能,从而影响精子发生。已有的研究结果表明,热激对支持细胞的分泌功能、超微结构、表达的骨架和连接蛋白、上游的核受体和细胞因子和血-睾屏障的通透性都产生可逆性的影响。
温度对支持细胞功能的影响
早期研究发现,隐睾或43℃局部热浴睾丸,可使雄激素结合蛋白(androgen-binding protein,ABP)和抑制素的生成和分泌明显减少;FSHR表达明显下降;胰岛素样生长因子1(insulin-like growth factor 1,IGF-1)表达增高。
中间纤维是支持细胞骨架的重要组分,正常成年动物支持细胞中的中间纤维主要是波形蛋白(vimentin)。在大多数成年隐睾患者的支持细胞中,中间纤维除波形蛋白外,结蛋白(desmin)和角质纤维蛋白(cytokeratin)也明显表达,而此后两者通常只在胚胎期和未成年支持细胞中表达。
角质纤维蛋白18可作为鉴定支持细胞成熟状态的标志分子,他仅表达于未成熟的支持细胞,而在成年睾丸的支持细胞不表达。成年猴隐睾手术或43℃水浴处理成年猴睾丸,可诱导其支持细胞重新表达CK18。停止热浴后,他的表达逐渐恢复到热浴前的水平(下图)。
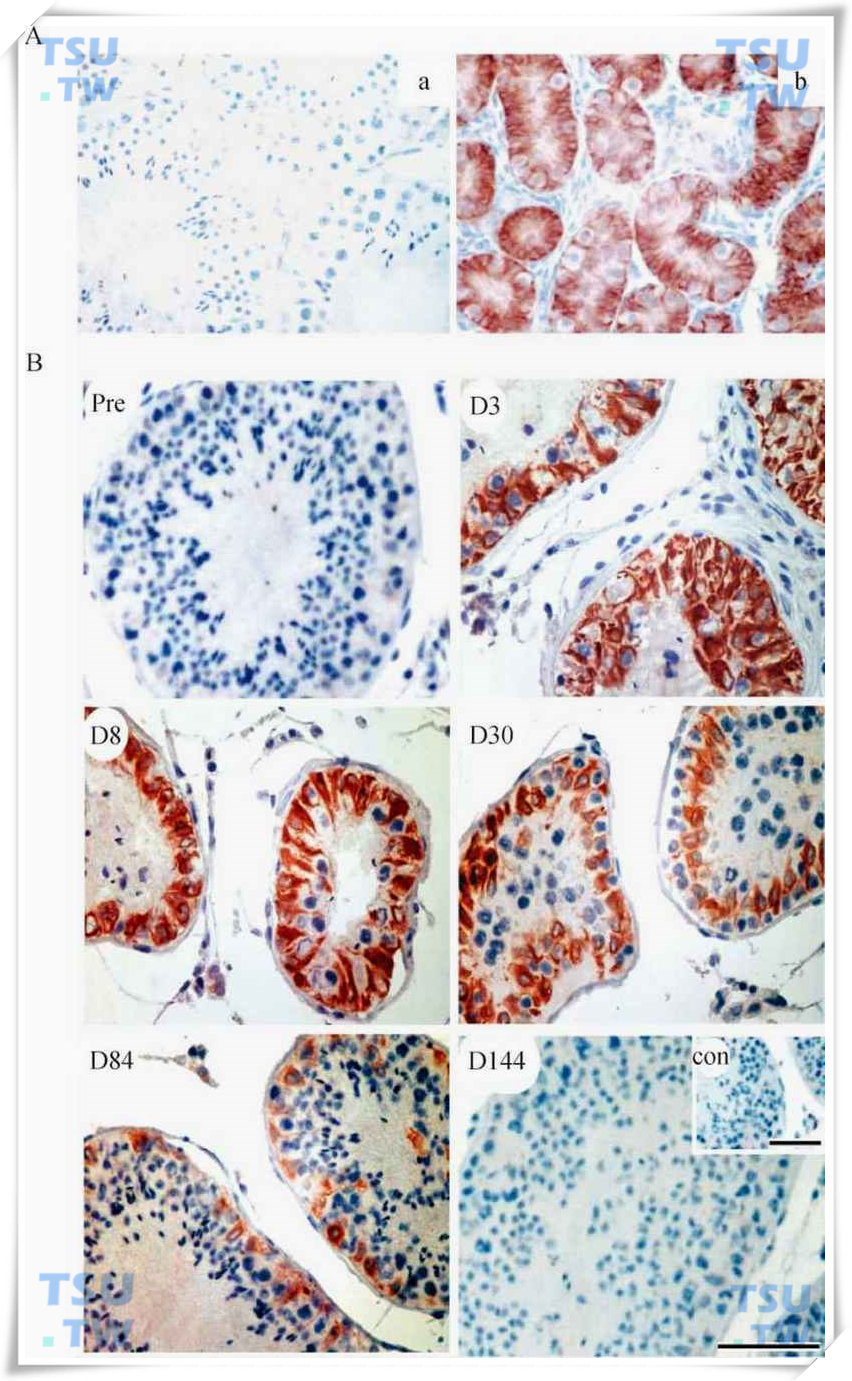
CK-18在猴睾丸中的免疫定位
A:CK-18在成年猴睾丸(a)和幼猴睾丸(b)中的免疫定位,显示CK-18仅在未成熟的支持细胞中表达。B:成年猴睾丸43℃热处理前及处理后第3,8,30,84,144天CK-18的免疫组织化学染色结果。红色为CK-18在支持细胞中的阳性信号,蓝色为背景颜色,con为阴性对照。标尺=100μm。(引自:Zhang XS et al.Endocrinology,2006,147(3):1237-1245)
肝细胞受体类似物(liver receptor homologue 1,LRH-1)是一个核转录因子,他在成年生精细胞、间质细胞和未成年支持细胞中表达,但不在成年猴或大鼠支持细胞中表达。与CK-18类似,在热处理成年猴或大鼠睾丸后,在分化的支持细胞中重新表达LRH-1,且其出现和消失的时间都比CK-18早。原代培养的成年猴或大鼠支持细胞的体外实验结果与在体实验结果一致(下图)。且ERK通路抑制剂可以阻止热激后CK18和LRH-1的表达。进一步的实验证实,LRH-1可能是调控支持细胞去分化的重要转录因子之一。这些研究结果提示,热激可能促使成年的支持细胞经历去分化过程,获得了未成熟支持细胞的特性。去分化的支持细胞丧失了支持精子发生的正常功能,从而影响精子发生。热激诱导支持细胞去分化,那么去分化的细胞是否具有增殖能力?在体和离体实验的结果表明,不论是隐睾还是43℃温浴,支持细胞中都没有Ki67的表达,表明虽然支持细胞发生了去分化,但是仍然不具有增殖特性,是一种病理的状态。
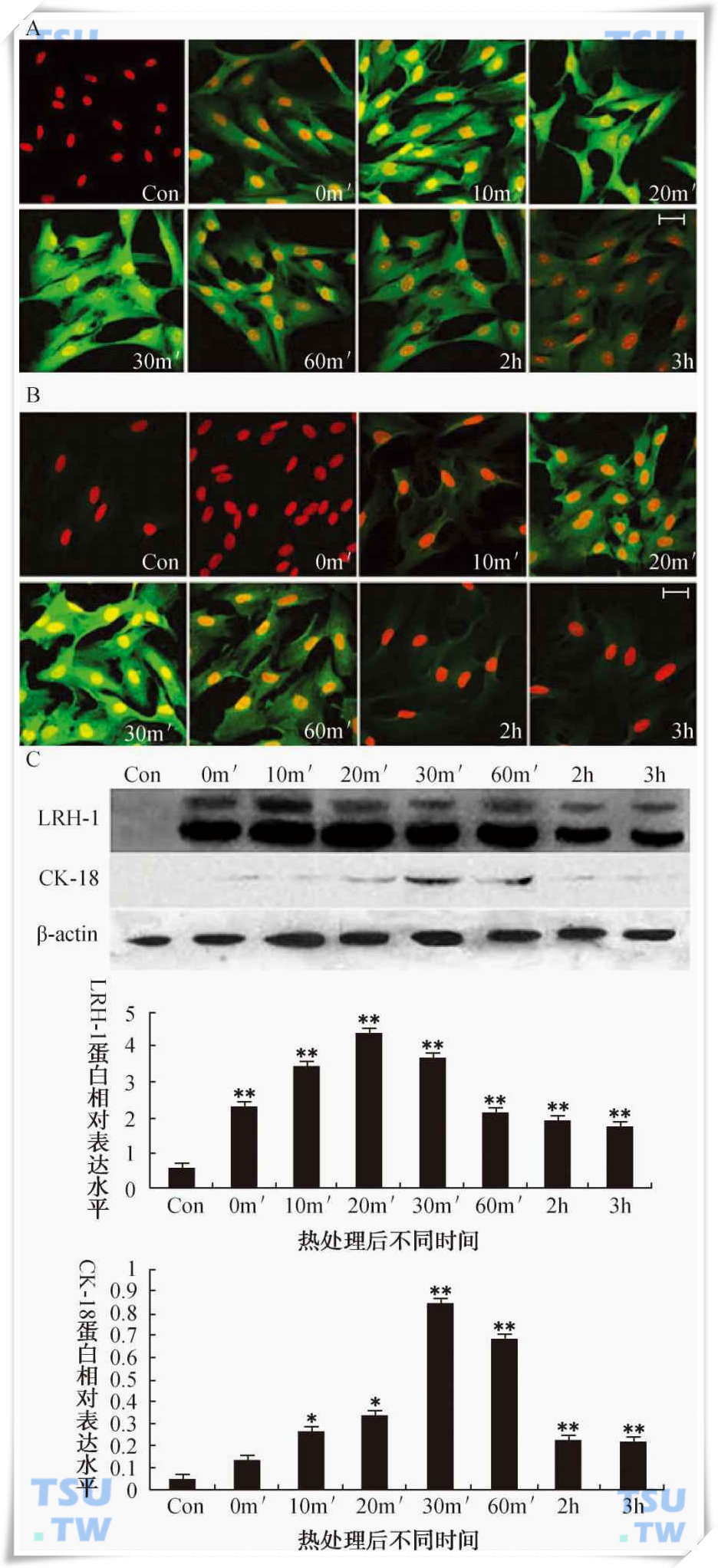
43℃水浴对LRH-1和CK-18在原代培养大鼠支持细胞中表达的影响
热处理后不同时间LRH-1(A)和CK-18(B)在支持细胞中的免疫定位。细胞核用PI标记。图中标尺=50μm。(C)LRH-1和CK-18的Western blot检测结果。相对表达水平为目的蛋白条带与内参β-actin条带的灰度比值,图中显示的数值为mean±SEM(n=3)。*代表与对照组相比P<0.05,**代表与对照组相比P<0.01。支持细胞分离自60日龄大鼠睾丸。0min、10min、20min、30min、60min、2h、3h表示43℃处理15分钟后不同时间。(引自:Guo Jian et al.Endocrinology,2007,148(3):1255-65)
热激可逆性改变生精小管内超微结构、增强血-睾屏障通透性
电镜发现隐睾患者或手术诱导隐睾大鼠的支持细胞结构上出现明显变化:内质网扩张,线粒体嵴明显减少,脂滴积累,电子致密物增加,核膜不规则,胞质空泡化,支持细胞中有各种不同直径的空泡积聚在细胞基底部,似乎是相邻细胞间连接处的细胞间隙局部扩大所致。四周后这种空泡消失。43℃热浴对生精小管内超微结构的变化更明显。热浴后6h已出现较大变化,在小管基膜处结构变得松散,可见较大空泡。支持细胞与生精细胞之间的连接处间隙明显变大。24h后这种现象更明显,胞质中可见许多空泡,这种空泡还出现在相邻支持细胞之间,使血-睾屏障出现断裂,但这种结构上的变化是暂时的,热浴后逐步恢复到正常状态。
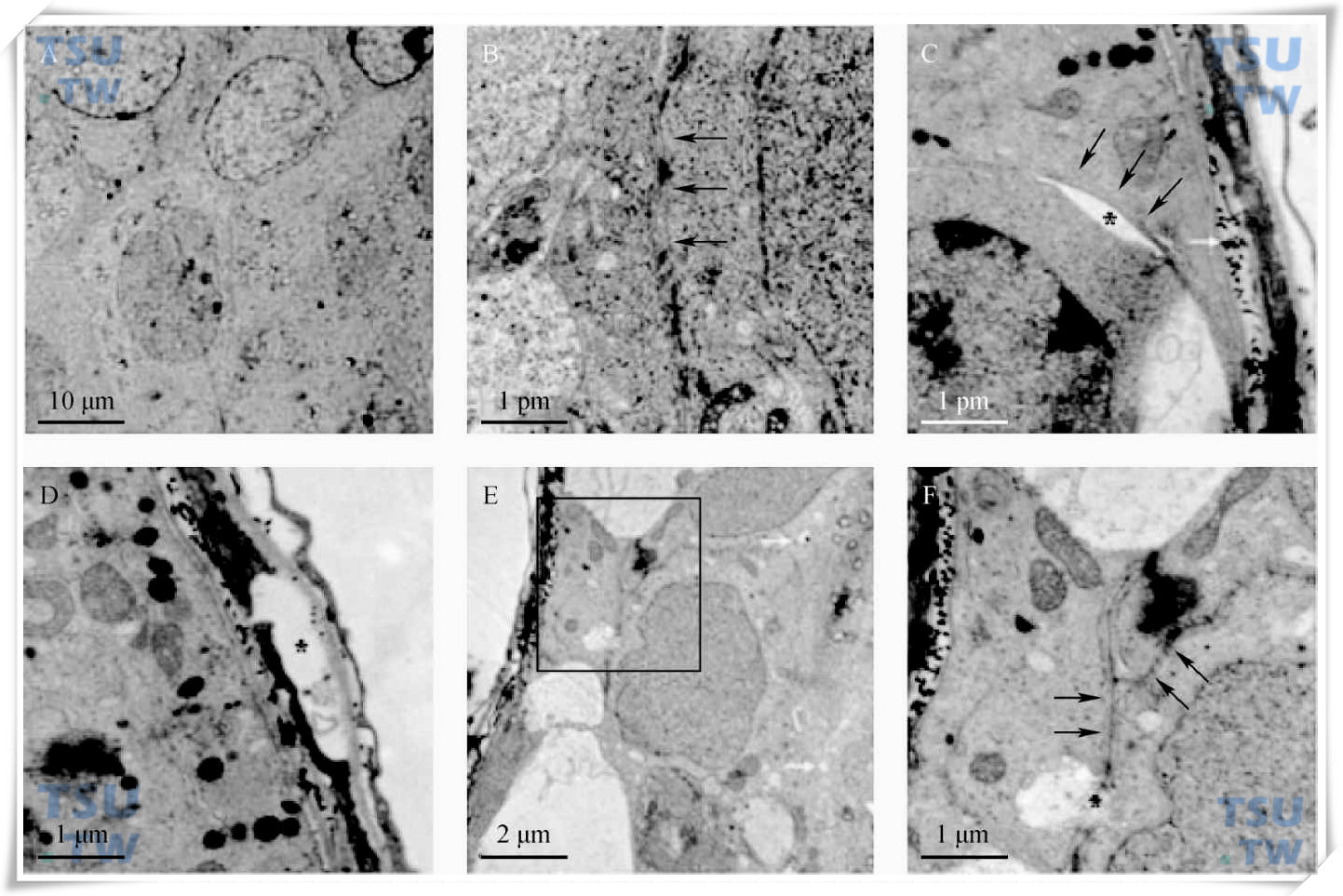
43℃水浴处理后大鼠睾丸曲细精管超微结构的变化
A和B:正常大鼠睾丸生精小管电镜图片。正常睾丸相邻支持细胞间的血-睾屏障是完整的(黑色箭头)。C和D:热处理后6小时。C:支持细胞和精原细胞间可见到细胞间隙(*),黑色箭头所指为锚定连接,白色箭头所指为基底膜内清晰可见的胶原。D:基底膜内可见较大空泡(*)。E和F:热处理后24小时。E:方框内为相邻支持细胞间的血睾屏障。此时支持细胞胞浆内出现更多的空泡(白色箭头)。F:E图方框内血睾屏障的放大图片。黑色箭头所指即为血睾屏障,此时血睾屏障处也出现了一些空泡(*)。放大倍数:2500(A);20 000(B,C,D,and F);8000(E)。(引自:Chen Min et al. Endocrinology,2008,149:4871-82)
生物素示踪被广泛用于检测血-睾屏障的功能,通过在睾丸白膜下生精小管间注射EZ-偶联的Sulfo-NHS-LC-Biotin可以检测血-睾屏障的通透性。向13d小鼠生精小管间注射的biotin能自由进入生精小管内,此时血-睾屏障还没有形成。出生18d左右,血-睾屏障开始形成,生精小管间注射的biotin可部分进入生精小管中,到成年后血-睾屏障完全形成,注射的biotin完全不能进入生精小管中。43℃水局部温浴成年小鼠睾丸30min,在热处理2d后生精小管管腔中发现有生物素存在,提示热浴破坏了血-睾屏障的通透性。但随着热处理后时间的延长,在热处理后10d左右,血-睾屏障通透性又恢复到正常水平(下图)。
43℃水温浴睾丸不同时间,对生精小管内精子发生和血-睾屏障通透性的影响不同。HE染色结果表明,热激30min或1h,10d后精子发生的恢复情况较好,血-睾屏障的通透性已经恢复到正常水平,而长时间热激(2h和3h)会明显地阻碍精子发生和血-睾屏障通透性恢复的时间。
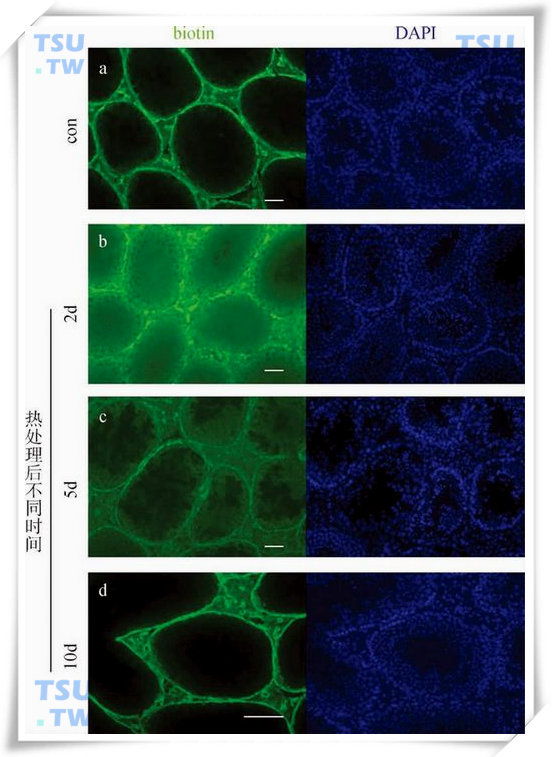
热处理可逆性增加成年小鼠血睾屏障通透性
热处理前后向小鼠睾丸白膜下注射EZ-Link Sulfo-NHS-LC-Biotin。a:正常组小鼠注射的生物素不能进入生精小管中。b:向热处理后2d的睾丸内注射生物素30分钟后,生物素渗透入生精小管中。c:热处理5d仍有部分生物素能渗透入生精小管中。d:热处理后10d血睾屏障通透性又恢复到正常水平。细胞核用DAPI标记,图中所有标尺=50μm。(引自:Cai Huan et al.Inter.J.Androl,2011,34(4):352-62)
热激对支持细胞骨架和连接分子表达的影响
在阴囊睾丸中,波形蛋白主要定位于支持细胞核的周围区域,偶尔呈纤维状伸向管腔。隐睾手术后波形蛋白除了表达量明显增强外,表达模式也发生明显改变,隐睾后10~15d,在支持细胞的整个细胞质中都能检测到波形蛋白的弥散表达。
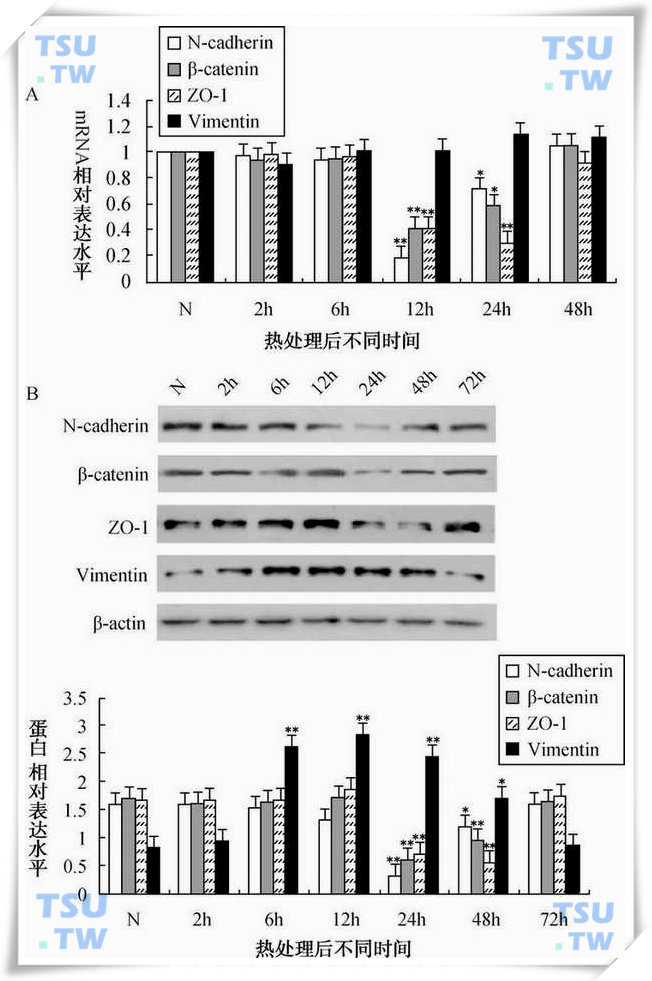
43℃热处理支持细胞后不同时间点N-cadherin,β-catenin,ZO-1和vimentin的mRNA及蛋白表达水平分析
A:实时定量PCR检测结果。N表示未处理组,2h,6h,12h,24h和48h分别表示热处理完成后不同时间。各处理组相对mRNA浓度以2-ΔΔCT计算,对照组浓度为1;B:Western blot检测结果。相对表达水平为目的蛋白条带与内参β-actin条带的灰度比值。图中显示的数值为mean±SEM(n=3).*代表与对照组相比P<0.05,**代表与对照组相比P<0.01。(引自:Chen Min et al.Endocrinology,2008,149:4871-82)
43℃水浴处理原代恒河猴支持细胞30min,热处理后将细胞重新放入33℃培养箱中继续培养。热处理后支持细胞胞质向核周回缩,胞体变圆,这种变化在热处理后2h最明显。6h后细胞形态又逐渐恢复到热浴处理前水平。ZO-1(zonula occludens-1)是构成紧密连接的重要蛋白,缺失ZO-1的小鼠不能形成紧密连接。N-cadherin和β-catenin是构成黏附连接的重要蛋白复合体,表达于支持细胞之间和支持细胞与生精细胞之间。黏附连接分子N-cadehrin、βcatenin和紧密连接分子ZO-1在热处理后24 和48h蛋白表达水平明显降低。他们的mRNA变化比蛋白的变化在时间上早12~24h。但中间纤维vimentin蛋白在热处理后表达明显升高,6h达到正常表达水平的3倍。在热处理后72h上述分子的表达水平逐步恢复到未处理前的细胞正常表达水平(上图)。小鼠的体内实验结果与此一致(下图)。由此可见,热激能可逆性地引起睾丸内黏附链接和紧密连接分子表达量和定位的变化,进而对连接的完整性和功能产生影响。
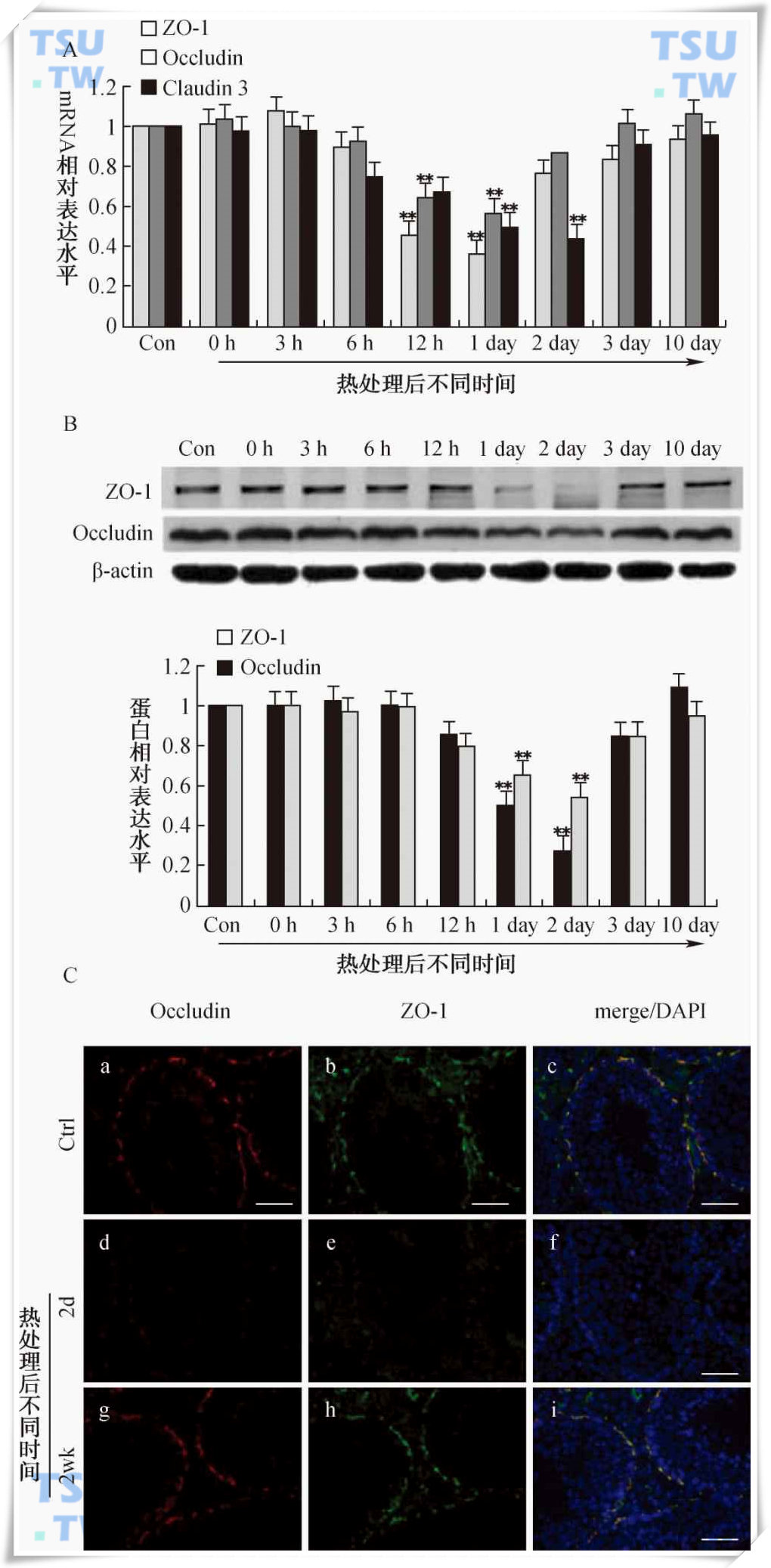
43℃热处理小鼠睾丸对紧密连接分子Occludin和ZO-1表达水平的影响
A:实时定量PCR检测结果。Con表示未处理组,0h、3h、6h、12h、1day、2day、3day和10day分别表示热处理(43℃30分钟)完成后不同时间。各处理组相对mRNA浓度以2-ΔΔCT计算,对照组浓度为1;B:Western blot检测结果。相对表达水平为目的蛋白条带与内参β-actin条带的灰度比值。图中显示的数值为mean±SEM(n=3),*代表与对照组相比P<0.05,**代表与对照组相比P<0.01;C:Occludin和ZO-1在睾丸中的免疫荧光分析。a,d,g中红色荧光为occludin的定位分布;b,e,h中绿色荧光为ZO-1的定位分布;c,f,i中蓝色荧光为DAPI染的细胞核,黄色荧光为红色和绿色荧光的叠加。其中a,b,c为未处理组睾丸;d,e,f为热处理后2天睾丸;g,h,i为热处理后2周睾丸。图中所有标尺=50μm。(引自:Cai Huan et al.Inter.J..Androl,2011,34(4):352-62)
热激对支持细胞上游转录因子和细胞因子的作用
雄激素和卵泡刺激素是雄性生殖中最重要的两种激素,他们对于支持细胞的增殖分化、精子发生的起始和维持都是不可缺少的。生精小管内,生精细胞并不表达雄激素和卵泡刺激素受体AR和FSHR,只有支持细胞表达这两种激素受体,雄激素和促卵泡生成素是通过支持细胞对精子发生起作用的。此外,WT1在胚胎性腺形成中是极其关键的转录因子,他在出生后及成年期的精子发生过程中同样起着关键的作用。恒河猴支持细胞的离体实验表明,FSHR在支持细胞中的表达在热处理早期没有明显变化,其表达的下调开始出现在热处理后12h,48h降至最低水平,72h恢复到正常水平。而与之相比,热处理15~30min后AR和WT1蛋白开始消失,6h后表达逐渐开始恢复。但其mRNA并未消失,只是表达明显减少。
睾酮和蛋白合成抑制剂放线菌酮并不能阻止这两个蛋白在热处理后的消失,26S蛋白酶体抑制剂也只有微弱的阻止作用。因此,这两个蛋白热激后消失的机制还有待于后续的研究。手术诱导大鼠隐睾的在体实验也发现,支持细胞中的AR和WT1在手术后也出现了可逆地消失,只是时相上慢一些,术后24h表达减弱,3d完全消失,10d恢复到正常水平,此时生精细胞的凋亡和脱落已非常明显。在时间上,AR和WT1的变化早于生精细胞的凋亡。在成人隐睾睾丸中,支持细胞表达AR的强弱可反映所在生精小管发育的成熟程度。在存在部分分化(尽管分化不完全)的生精细胞的小管,支持细胞中AR表达强;在无生精细胞的小管,支持细胞中AR表达微弱或没有表达。由此可见,支持细胞中AR表达的缺失与小管局部精子发生缺失相关。因此可以认为,AR和WT1的消失是43℃水浴或手术隐睾引起生精细胞凋亡的重要原因之一。
此外,雄激素拮抗剂flutamide对支持细胞中骨架和连接分子的作用与43℃水浴的作用基本相似,即抑制N-cadherin、β-catenin、ZO-1等连接分子的表达,上调中间纤维vimentin的水平。腺病毒载体介导的外源AR能补救热处理后内源AR蛋白的消失,过表达AR能挽救热处理引起的骨架和连接分子的变化,表明热处理后支持细胞中AR蛋白的消失介导了随后骨架和连接分子的变化,但是对于其分子机制尚不清楚。文献报道,转化生长因子β(transforming growth factorβ,TGF-β)以一种时间特异性方式由睾丸支持细胞和生殖细胞所分泌,他们可以在转录水平、转录后水平和翻译后水平调控细胞连接的重构。热激可引起睾丸内TGF-β3表达可逆性增高,伴随热激向睾丸内注射TGF-β拮抗剂可以部分阻止热诱导的血-睾屏障的破坏,提示TGF-β3可能是热激引起紧密连接破坏的一个调节因子之一(下图)。
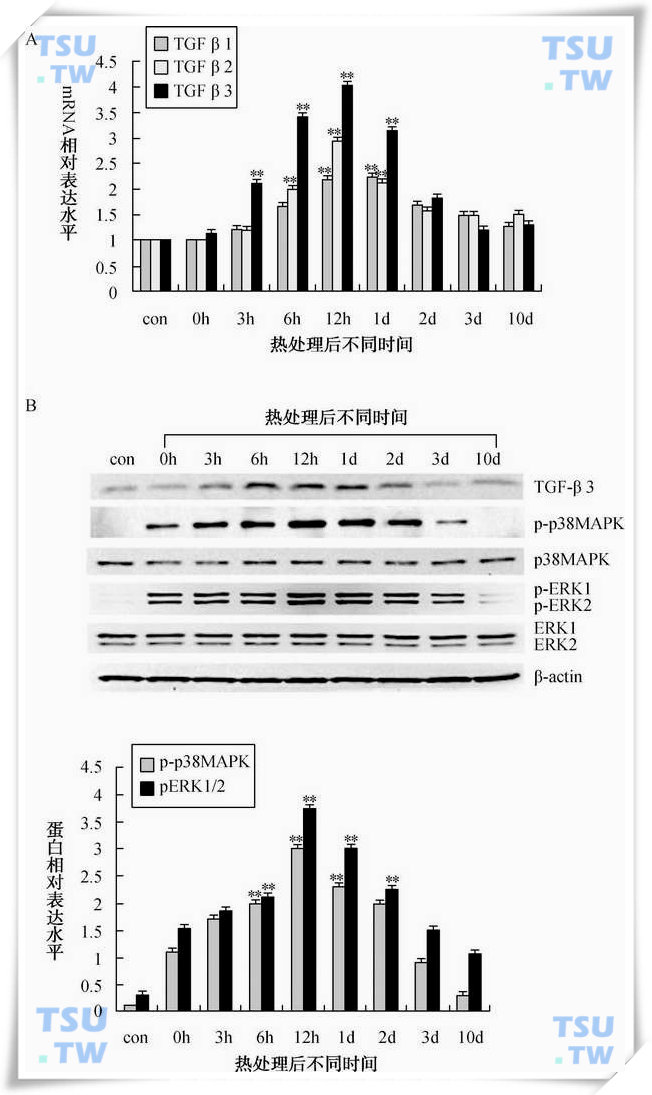
热处理诱导小鼠睾丸TGF-βs表达并活化p38MAPK和erkMAPK
A:实时定量PCR检测TGFβ1,2,3的mRNA在热处理后小鼠睾丸中的表达变化。各处理组相对mRNA浓度以2-ΔΔCT计算,对照组浓度为1。B:Western blot检测热处理后小鼠睾丸内TGF-β3和p38MAPK、erkMAPK磷酸化表达变化。p-p38MAPK为磷酸化p38MAPK,p38MAPK代表总的p38MAPK水平。p-ERK1/2为磷酸化ERK1/2MAPK,ERK1/2代表总的ERK1/ 2MAPK水平。相对表达水平为磷酸化目的蛋白条带与总蛋白条带的灰度相对比值,对照组设为1。图中显示的数值为mean±SEM(n=3),*代表与对照组相比P<0.05,**代表与对照组相比P<0.01;con表示未处理组,0h、3h、6h、12h、1d、2d、3d和10d分别表示热处理(43℃30分钟)完成后不同时间。(引自:Cai Huan et al.Inter.J..Androl,2011,34(4):352-62)
AR和WT1是生精过程必不可少的两个因子,他们各自的支持细胞条件敲除小鼠其精子发生均受到明显的抑制。作为上游因子,AR和WT1在支持细胞中的作用肯定不仅仅是调节细胞连接,热处理后他们的表达变化给支持细胞带来的其他影响和机制还有待挖掘。
综上所述,热激可诱发支持细胞发生去分化现象,支持细胞热激后虽然未发生凋亡,但他的细胞极性、分化状态、分泌活性、超微结构、表达的上游核受体和细胞因子以及骨架连接分子都受到了显著而可逆的影响。生精小管中支持细胞与生精细胞紧密相关,精子发生的正常进行有赖于支持细胞提供营养物质和结构的支持。热激后支持细胞结构和功能发生显著变化,必然干扰了支持细胞对生精细胞的支持作用,这可能是热激诱发生精细胞凋亡的一个重要机制。

热激影响精子发生的作用途径
