
腺垂体可合成和分泌生长激素(growth hormone,GH)、泌乳素(prolactin,PRL)、促甲状腺素(thyrotropin,TSH)、促肾上腺皮质激素(adrenocorticotropic hormone,ACTH)、黄体生成素(luteinizing hormone,LH)、促卵泡激素(follicle-stimulating hormone,FSH)等10多种激素。
GH分子形式不均一
人GH基因位于第17号染色体长臂(17q22-24),由5个同源性很高的基因组成,全长约66kb,包括5个外显子和4个内含子。循环血中的GH分子形式是非均一性的,包括多种形式的单体、同源或异源聚合体、分子片段和单体与其结合蛋白的复合体等。其主要形式是分子量为22kD的GH单体,等电点5.1,由191个氨基酸残基组成,含4个半胱氨酸,形成2个分子内双硫键,使分子具有大小两个环(下图)。22kD的GH约占垂体GH的70%~75%,占循环血GH的43%。其次是20kD的GH单体(含176个氨基酸残基,缺乏第32~46位15个氨基酸残基)。20kD的GH占垂体和循环中GH的5%~10%。第3类GH单体分子是酸性GH。
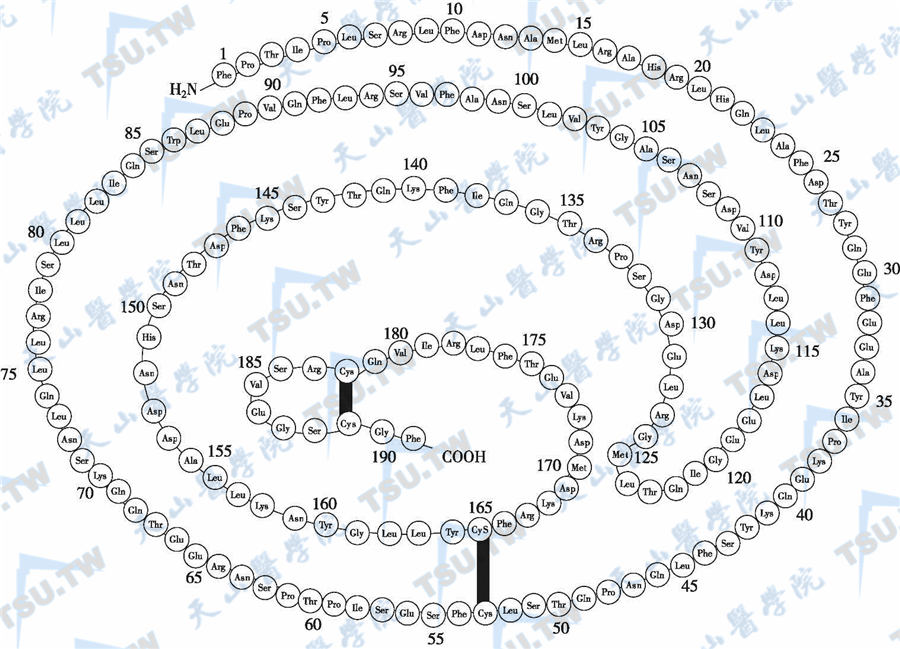
垂体生长激素结构图
GH是机体生长与代谢的调节因子
GH能促进骨、软骨、肌肉以及其他组织的细胞分裂、增殖和蛋白质合成。GH诱导靶细胞产生生长介质(somatomedin,SM),即胰岛素样生长因子-1(insulin-like growth factor-1,IGF-1)和IGF-2。GH的促生长作用主要通过IGF-1介导。GH促进蛋白质合成,增强钠、钾、钙、磷、硫等的摄取和利用,抑制糖消耗,加速脂肪分解,使机体的能量来源由糖代谢向脂肪代谢转移,有利于生长发育和组织修复。
GH的促合成代谢作用是通过刺激IGF-1、胰岛素和游离脂肪酸的作用实现的。当患者的营养状况正常时,GH引起的IGF-I和胰岛素是合成储存物质及瘦体重(lean body mass, LBM)增长、脂肪和糖原合成的关键因素。在基础状态下,GH促进蛋白合成,降低其分解,但在禁食和应激状态下,缺乏GH可加剧蛋白质的丢失(约50%)。GH拮抗胰岛素的作用,诱导胰岛素抵抗,并成为应激和感染时高血糖症(应激性高血糖症)和黎明现象(dawn phenomenon)的病理生理基础。
GH受体以激酶交联方式作用于靶细胞
人GH受体(GHR)基因编码638个氨基酸,其中包括18个氨基酸残基组成的信号肽。GHR分子为单肽链糖蛋白,其N端216个氨基酸残基中含有5个潜在的糖基化位点,位于细胞膜外侧,构成GH结合结构域;第247~270个氨基酸残基链为强疏水性穿膜肽;C端350个氨基酸位于胞质膜内侧,构成信号转导结构域。GHR的实际分子量>100kD,与cDNA推导值(70kD)差异悬殊,其原因是GHR有众多糖基和泛肽结构。机体许多组织细胞都表达GHR,如肝、脑、骨骼肌、心、肾、肺、胃、肠、软骨、胰腺、睾丸、前列腺、卵巢、子宫、脂肪细胞、成纤维细胞和淋巴细胞等。GH与其受体结合,直接或间接促进生长发育。在胎儿和新生儿期,各类细胞上的GHR数量最多,对GH的反应十分敏感。
GH与细胞膜上的受体结合,刺激受体以及多种细胞内的蛋白酪氨酸磷酸化。奇怪的是,GH/PRL/细胞因子受体家族成员的胞内部分均不存在同源序列,但能分享共同的信号途径。配体结合胞膜受体,诱导二聚化,激活胞内JAK-2,并进一步将信号向下游传导。JAK激酶的激活出现胞内多种蛋白质磷酸化,主要有JAK-stat途径、ERK1和ERK2(MAP激酶途径)、S6激酶途径等。DAG/PKC还参与了GH的胰岛素样效应途径,G蛋白、磷脂酶C参与了GH的胰岛素抵抗效应及降脂效应。GH与GHR结合后,可经细胞内吞作用进入胞质并可越过核膜进入核内直接发挥作用,核染色质上存在GH结合蛋白(GH binding protein,GHBP)或GH的结合位点,GH的核转位不依赖于细胞骨架系统。
GH分泌受多种因素调节
GH释放激素
下丘脑GH释放激素(GHRH)是腺垂体分泌和合成GH最主要的生理性刺激物。GHRH的分泌是暴发脉冲式的。GHRH刺激GH的分泌、合成和GH细胞增生。
生长抑素
下丘脑神经元合成的生长抑素(somatostatin,SS)经轴突输送到正中隆突,进入垂体门脉血管,到达腺垂体,不但抑制GH的基础分泌,而且抑制GH对生理性和药理性刺激,如运动、精氨酸与胰岛素低血糖和GHRH等引起的兴奋反应。SS也能抑制垂体GH瘤、糖尿病及Laron矮小症患者异常升高的血清GH水平。
GHRH和SS是所有到达下丘脑并影响GH分泌的最后整合通路,GH分泌是这两种下丘脑激素相互拮抗作用平衡的结果。自发性GH分泌峰是由于大量GHRH释放和低水平SS作用的结果。SS可直接抑制下丘脑释放GHRH,也可通过超短反馈抑制GHRH神经元的活动。GHRH神经元可兴奋SS神经元,SS与GHRH神经元之间的交通是双向的,两种神经元之间有突触联系。
葛瑞林
葛瑞林(ghrelin)是GHRH、食欲刺激肽和GH释放激素受体(GH releasing hormone receptor,GHSR)的内源性配体,由28个氨基酸残基组成。体内存在两种形式的葛瑞林,一种是无活性的N端去辛酰基(capryloyl);另一种是有活性的N端辛酰基,即第3位丝氨酸的N端18-辛酸以酯键连接。葛瑞林主要由胃分泌,但在肠、胰腺、肾、免疫系统、胎盘、睾丸、脑垂体和下丘脑等组织中都有低度表达,通过促GH分泌素(GHS)受体1a(GHS2R)而发挥作用。葛瑞林受体主要分布在下丘脑、腺垂体和胃,肾和卵巢中也有少量分布。葛瑞林是强效的促进GH释放物质,能促进体外培养的垂体细胞释放GH。给人静脉注射葛瑞林,引起剂量依赖性GH释放,但不能刺激GH合成。对糖代谢和脂代谢有影响,同时还影响食欲、胃肠活动、睡眠、记忆、行为、免疫、细胞增殖和生殖功能。
神经递质
在GH分泌中起重要作用的神经递质有:①多巴胺(DA)刺激GH分泌。②去甲肾上腺素对GH具有兴奋和抑制的双重作用,α2受体激动剂可乐宁可刺激GH分泌。α2受体阻断剂抑制或减弱人对精氨酸、低血糖、运动、电休克等各种刺激引起的GH分泌,但不影响GH的基础释放及脉冲性分泌和与睡眠有关的GH分泌。另一方面,去甲肾上腺素对GH分泌又有抑制作用,β受体阻断剂加强GH 对GHRH的反应。③乙酰胆碱(acetylcholine)对GH有兴奋作用。阿托品则抑制L-多巴、精氨酸、胰高血糖素、可乐定(clonidine,可乐宁)、运动等刺激及与睡眠有关的GH分泌。④阿片肽、5-羟色胺和γ-氨基丁酸对GH分泌有兴奋和抑制两种作用,可能取决于刺激的强度和干预条件。组胺H2受体阻断减弱L-多巴和低血糖兴奋GH分泌的作用。此外,胆囊收缩素、VIP、阿片肽等在中枢神经系统中参与了GH分泌的调节。
垂体旁分泌激素
GH细胞分散分布于腺垂体的大部分,一些细胞沿血管分布,常与ACTH细胞并行排列,常被ACTH细胞的胞突包围;还有一些GH细胞与TSH细胞密切联系,并被滤泡星状细胞环抱。腺垂体细胞还能合成DA、5-HT和GABA等,调节GH细胞的功能;活化素(activin)减弱基础和GHRH兴奋的GH分泌,抑制GH转录;AT-2减弱GH对GHRH的反应。
其他激素
可能有多种:①甲状腺激素过多或缺乏导致GH释放迟钝,其机制可能与调节生长抑素(somatostatin,SS)和GHRH的作用有关;②糖皮质激素抑制GHRH引起的GH释放;③雌激素对GH分泌有易化作用,因此妇女比男性有更高、更频的自发GH分泌峰,并对经典的GH刺激试验有更强的反应。
