
胚胎发育过程中血管内皮细胞和造血细胞均来源于中胚层,长期以来就推测血管系统和造血系统存在共同的前体细胞,即血液血管干细胞。最近的研究结果也支持该假说:①以前认为的几种造血系统特异性因子和白细胞介素家族如G- CSF、GMCSF、IL- 3、IL-4、IL-6、IL- 8均证实对内皮细胞功能有一定的影响。在IL- 1或TNF刺激下,内皮细胞也能产生某些调节造血的因子如IL- 1、IL-4、IL-6、GMCSF等。②内皮细胞表面表达某些造血细胞的表面抗原,如鹌鹑QH1和MB1以及人或鼠的CD34、CD31。③人类内皮细胞也表达和造血调控相关的IL- 3、kit配体、EPO和TPO的受体。
内皮细胞和巨核细胞表面抗原的相似性以及调节两类细胞发育的因子间的相互重叠进一步证实了内皮细胞和造血细胞存在共同的起源。目前研究显示内皮细胞是支持造血生成,特别是巨核细胞生成的骨髓基质的主要成分,体外共培养骨髓内皮细胞(BMEC)和造血祖细胞显示:与其他种系的造血细胞相比,巨核细胞的增生更为显著。体外实验已证实BMEC通过分泌kit配体、IL-6、TPO等因子支持巨核系祖细胞的增殖和分化,由此可以推测在体内BMEC对巨核细胞的生长和成熟起一定的作用。此外,在缺乏外源性内皮生长因子的情况下,巨核细胞的增殖伴随着内皮单层的长久维持,这表明巨核细胞也可能产生有助于内皮细胞存活和增殖的因子。
血管新生,即从已存在的血管的基础上形成新的血管,存在于某些正常和病理情况下,如伤口愈合、肿瘤生长等。血管新生是一个复杂的多步骤过程,反映了复杂的正、负血管新生调节物的消长平衡,作为各种血管性调节因子作用的靶细胞,内皮细胞在血管新生中发挥了重要的作用,其中某些血管性调节因子也调节巨核系细胞生成。
以下描述内皮细胞和巨核细胞的共同抗原和共同的调节因子,以及这些调节因子对血管新生和巨核细胞生成的影响,从而进一步探讨两者共同的起源。
巨核细胞和内皮细胞共同表达的抗原
巨核细胞和内皮细胞合成和表达在生物学上和免疫学上相同或相似的膜糖蛋白、胞质因子和胞内小体,以及都表达VEGF和FGF的受体。
巨核细胞和内皮细胞共同表达的抗原

巨核细胞和内皮细胞的共同调节因子
巨核细胞和内皮细胞的共同调节因子
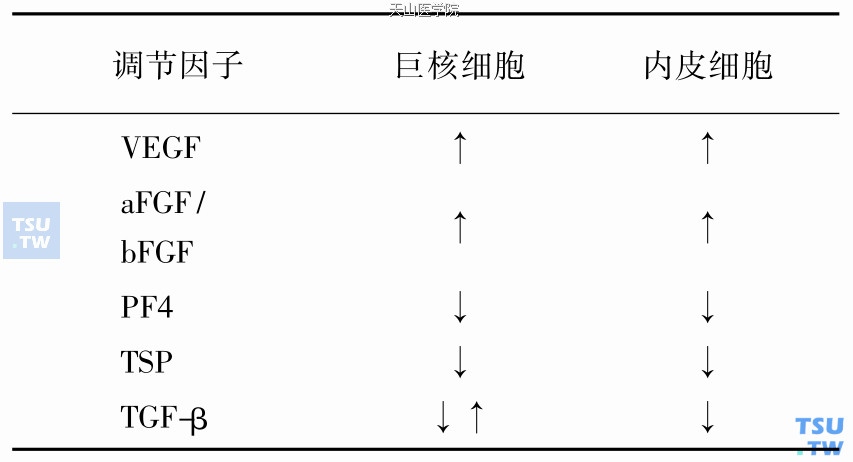
VEGF和VEGF受体(KDR)
VEGF是一种特异性的内皮细胞分裂原,能影响内皮细胞的形状改变和迁移,增高血管通透性,在体内外都有强烈的刺激血管新生的作用。在氨基酸水平上与血小板来源的生长因子家族如VEGF- B、VEGF- C、PDGF有15%~25%的同源性。
VEGF的活性形式为同源二聚体,分子量为34~46kDa,不同的外显子选择性剪接产生大小不同的4种形式:121、165、189和206个氨基酸,后三种具有肝素结合位点。VEGF- 121和VEGF- 165为分泌型,而VEGF- 189和VEGF- 206存在于膜表面或沉积于细胞外基质。在斑马鱼(zebrafish)的胚胎中,VEGF能同时刺激造血细胞和内皮细胞的分化,而且共同表达Flk- 1受体。正常的细胞和转化的细胞都可以产生VEGF,在许多存在血管新生的生理和病理状况下,VEGF过度表达。在存在血管新生的肿瘤中,VEGF表达上调,可能是因为癌基因的激活或肿瘤抑制物的丢失。现在认为VEGF既可促进血管新生,也可促进血管生成,即在出生后类似于胚胎发育过程中的从血管前体细胞分化形成新的血管。Chris- toph等通过给冠心病患者心肌注射携带有VEGF- 165 DNA片段的质粒(phVEGF- 165),引起外周血VEGF水平的一过性增高,并且动员骨髓来源的内皮前体细胞释放到外周血,在一定部位分化成成熟的内皮细胞,形成血管,提高缺血心肌的灌注量。
Osamu等在CD34阳性细胞、巨核细胞和血小板中通过RT- PCR检测到VEGF的受体KDR和Flt-1的mRNA,与免疫组化结果显示在这几类细胞中存在VEGF蛋白相符合。已知造血细胞能产生VEGF- 121、VEGF- 165,可以推测VEGF可能调节造血干细胞和巨核细胞的生存、生长和分化,同时造血细胞产生的VEGF也能够分泌到骨髓微环境中调节BMEC的功能。研究发现IL- 3、TPO在体外刺激巨核细胞或凝血酶诱导的血小板聚集能引起VEGF的释放。在体内骨髓巨核细胞增殖状况下,血小板能够储存和运输VEGF到血管损伤部位。可见在巨核细胞和血小板系统,VEGF具有以下的生理功能:①在骨髓微环境中,VEGF作为一种旁分泌因子调节巨核细胞的增殖和分化,即增殖的巨核细胞分泌VEGF和其他造血因子刺激自身的生长、黏附以及最后的成熟和血小板的释放。②VEGF是一种重要的损伤修复因子,在与伤口修复有关的血小板聚集、血栓形成和血管新生等过程中均起作用。
VEGF通过结合两种酪氨酸激酶受体Flt- 1和KDR(小鼠中为Flk- 1)而起作用,现在认为KDR在VEGF功能的信号传导中起关键作用,刺激内皮细胞的DNA合成,与VEGF的增殖活性密切相关。KDR是第一个在血液血管干细胞表面表达的内皮细胞酪氨酸受体,小鼠Flk- 1靶基因敲除实验显示Flk- 1对于血液淋巴细胞生成和血管形成是必不可少的。出生后内皮细胞表达KDR,在造血祖细胞和巨核细胞中存在KDR的mRNA,但在其他成熟造血细胞中没有发现。可见,VEGF- KDR信号传导途径是巨核细胞生成调节和血管性因子调节血管新生的因子网络的一个重要连接点,反映了内皮细胞系和巨核细胞系的相互作用。
血管生成素(angiopoietin)作为调节血管生成的另一类酪氨酸受体家族TIE/TIE1和TIE2/TEK的配体,在体内能促进VEGF诱导的血管新生,在体外协同VEGF促进内皮细胞和造血细胞的增殖。Suda等体外培养造血功能缺失的AML缺陷性胚胎的前AGM区,该胚胎的头部、心包和胎肝存在缺陷性的血管新生,通过与表达血管生成素的造血干细胞共培养,直接促进了内皮细胞的迁移,受损的血管新生也获得改善。由此可见,造血干细胞对于胚胎形成时期的血管新生是必需的。
FGF
aFGF和bFGF是肝素结合生长因子家族的主要成员,能够刺激多种细胞的生长和分化。已经发现4种不同的FGF受体家族:flg(FGFR1)、bek(FGFR2)、FGFR3和FGFR4。FGF在体内外均刺激内皮细胞生长,引起血管新生。同样FGF也在不同程度影响造血生成,可能是直接作用于干细胞及各系祖细胞,也可能与基质细胞相互作用,间接引起造血因子的释放,从而刺激造血生成。已知aFGF 和bFGF是巨核细胞生成的正性调节因子,两者在体外均能刺激巨核祖细胞和HEL细胞系的生长,体内给予FGF能够增加小鼠外周血血小板。其作用机制,FGF最可能是间接诱导骨髓巨核细胞或骨髓基质细胞分泌某些因子如IL-6。免疫荧光染色显示巨核细胞和血小板含有大量的FGF,同时巨核细胞也表达在内皮细胞上发现FGF的受体flg和bek。
PF4
PF4以非同源四聚体的形式储存在血小板,血小板激活时被释放出来。PF4属于C- X- C家族,能够黏附内皮细胞和巨核细胞,抑制血管新生和巨核细胞生成。加入外源性的肝素或与PF4肝素结合位点相似的合成肽段能阻止PF4黏附功能,从而阻止其对两类细胞的抑制作用,提示PF4可能通过相同的机制影响巨核细胞生成和血管新生。
已知bFGF和VEGF通过结合细胞特异性受体促进有丝分裂功能。在内皮和巨核细胞系都能检测到bFGF和VEGF受体。在与特异的受体结合前,Bfgf/VEGF和细胞表面的肝素样分子HSPG结合,后者作为低亲和力受体调节这两种受体和配体的结合,PF4可能通过其羧基末端的一个碱性氨基酸簇构成的主要肝素结合区竞争结合bFGF/VEGF,阻止bFGF/vEGF的内聚化和二聚化,从而破坏HSPG和FGF/VEGF相互作用,抑制细胞的生长。然而,长期以来观察到缺乏肝素亲和力的PF4类似物在体内仍具有抑制血管新生的能力,而含有PF4主要肝素结合区的P58~70片段在体内外实验中却不能干扰小鼠巨核细胞的生成,由此推测肝素结合位点并不是PF4抑制巨核细胞生成和血管新生的唯一因素。Han等发现PF4的另一条不依赖肝素的途径通过中心功能区而不是肝素结合序列来抑制造血,主要是抑制巨核细胞生成和血管新生,表明肝素结合位点并不对所有的血管新生都必要。通过化学因子结构功能相互关系的研究显示,位于7~9位和54~56位氨基酸的两个DLQ结构域(Asp- Leu- Gln)对于抑制髓系祖细胞增生的蛋白是必需的。因此,可以假设PF4首先与细胞表面的肝素类似因子相互作用引起该分子的构象改变,从而有助PF4的中心功能区与细胞接触,最终抑制此两类细胞的增生。
TSP- 1
TSP- 1是TSP家族的一员,为一个大分子量的(450kDa)同源三聚体的黏附性多糖蛋白,是血小板-α颗粒的主要成分,也存在于多种细胞,如内皮细胞、平滑肌细胞和成纤维细胞,在细胞激活时大量释放。TSP- 1含有多个功能区,其中肝素结合位点尤为重要。TSP- 1在细胞基质间和细胞间的相互作用方面十分重要,能够介导包括造血干细胞在内的许多细胞的黏附。目前研究显示,TSP- 1与许多血小板来源的因子如PF4、β-血小板球蛋白和TGF-β相似,全长及其肝素结合区片段能够通过调节造血生长因子如TPO和bFGF的活性从而抑制巨核细胞生成,而且肝素能中和这种抑制反应。
TSP- 1能抑制巨核细胞生成,也能抑制内皮细胞的形成,从而抗血管新生。完整的TSP- 1和含有肝素结合肽段的TSP- 1片段能抑制bFGF对内皮细胞刺激增生和迁徙的功能,也能抑制几种肿瘤动物模型或正常和病理状况的血管新生,其抗血管新生活性主要位于3个含60个残基的flg重复序列。因为结合HSPG是呈递bFGF到高亲和力的bFGF受体所必需的,可以推测竞争性结合bFGF的细胞表面的HSPG至少部分解释TSP- 1及其片段的血管新生的抑制作用。目前发现TSP- 1及采自flg重复肽段的肝素,其结合的特异性与bFGF相重叠。这些结果均表明TSP- 1作为巨核细胞生成和血管新生的负性调节因子对两者有相似的作用机制。
TGF-β
TGF-β属于多肽家族,且有与生长分化和炎症等相关的多种生物功能,其中对内皮细胞生长的抑制作用最为明显。然而,TGF-β对血管新生有双重效应:低浓度的TGF-β协同提高血管新生,而高浓度的TGF-β可降低VEGF或bFGF所诱导的体外内皮细胞的血管穿透性。虽然在体外TGF-β抑制其他促内皮细胞增生迁徙的血管因子的活性,但在体内能刺激血管新生,可能是因为TGF-β对各种细胞,如成纤维细胞和单核-巨噬细胞的趋化性不同,在组织中TGF-β通过不同的浓度梯度募集必需的结构细胞到血管新生位点。TGF-β作为血管性因子基因表达的强有力的刺激物对增加细胞分泌VEGF起主要作用,相对于单个细胞培养,这种效应在复杂组织中更为常见。在造血系统中,TGF-β是巨核细胞增生和多倍体化的强有力抑制物,抑制巨核细胞核内有丝分裂,对巨核细胞的作用呈现浓度和因子依赖性,表明TGF-β或者通过拮抗FGF和IL- 3的功能起抑制作用,或者通过影响炎性细胞间接起刺激作用,主要抑制早期的而非晚期的生长因子。体外培养的巨核细胞分泌TGF-β并不对巨核细胞的多倍体化起自分泌的调节作用。
ECM
除了可溶性因子,ECM也能以独立或协同的方式影响细胞的功能。在ECM分子中,蛋白多糖广泛地与细胞因子和生长因子协同作用,通过核心蛋白以及黏多糖链和细胞因子相互作用,调节细胞因子和基质细胞表面受体间的作用,甚至黏多糖自身也具有因子的活性。
肝素和硫酸肝素是两种重要的黏多糖,ECM通过它们作为细胞因子和肝素结合因子联系的纽带,例如硫酸肝素蛋白多糖调节bFGF和它的受体之间的相互作用,稳定配体受体复合物,保护bFGF不被降解以及有助于bFGF的二聚体化。在这些ECM结合的细胞因子中,bFGF、c- kit配体和GM- CSF是促进巨核细胞生成的正性调节因子,而TGF-β是强有力的负性调控因子。同时肝素和硫酸肝素也是非常重要的血管新生调节物,两者通过与细胞结合调节细胞对血管性因子FGF、VEGF、PF4和TSP- 1的反应。
从上述对巨核细胞和内皮细胞共同的抗原以及巨核细胞生成和血管新生的共同调节因子的总结,表明这两类细胞在发育过程的相似性和在功能上可相互作用。巨核细胞和内皮细胞很有可能来自同一前体细胞。进一步深入研究两者的细胞抗原和相关基因,以及影响两者发育的共同因子,可以增加我们对巨核细胞系和血管性疾病,尤其是涉及血管新生的病理机制以及两者之间相互关联的理解,有利于在未来针对这些疾病发展新的治疗策略。
(韩忠朝 陈芳)
