
血液中多种促凝蛋白是维生素K依赖性凝血蛋白。自20世纪50年代起维生素K拮抗药(双香豆素及华法林)一直用于治疗血栓病。r-羧基谷氨酸(Gla)转录后修饰过程的记叙(1974),不仅对理解凝血机制是一种突破,并且也促使发现了蛋白C通路,一种依赖维生素K的抗凝途径。从牛血浆中纯化获得蛋白C(PC)后不久,获知PC是一种维生素K依赖性血浆酶原蛋白(Stenflo 1976)。PC被凝血酶活化而具有抗凝活性(Kisiet等1977,1979)。APC与Mammen(1960)描述的自激凝血酶原Ⅱ- a (autoprothrombin Ⅱ- a)为同一物质(Seagers等1976)。已阐明自激凝血酶原Ⅱ- a虽可由凝血酶原制备工艺中加入凝血酶一同温培而产生,但并非来自凝血酶原的碎片(Maciniak 1972)。回顾分析认为该系统中可能存在PC的污染。PC抗凝过程分为三阶段:①PC必须在凝血酶与TM形成(T- TM)复合物的作用下才能被激活成为APC;②APC能使与膜结合着的、被凝血酶活化的凝血因子Ⅴ和Ⅷ(FⅤa、FⅧa)产生蛋白溶解性降解;③血浆蛋白酶抑制物对APC的抑制作用及在循环中被清除。
凝血酶自身是PC的弱激动剂。Owen和Esmon (1981)发现的TM是这一反应的辅因子。TM存在于内皮细胞表面,能以高亲和力和产生在其邻近地区的凝血酶相结合。T- TM复合物使凝血酶失活并产生激活蛋白质C的活力。循环系中唯独微循环系的毛细血管床具有最高的表面积和体积比。微循环中高水平的TM对PC的局部性活化和血液的抗凝作用至关重要。最近阐明的一种内皮细胞PC受体(endothelial cell protein C receptor,EPCR),能结合PC的Gla区、向活化的T- TM复合物呈递PC。EPCR对PC的活化作用极为重要。APC裂解凝血系的2个重要辅因子FⅤa和FⅧa的几个肽键而使凝血通路受抑制,因此能下调凝血酶的生成反应。APC在循环中的半衰期相对较长(约20分钟)。APC被PC抑制物(protein C inhibitor,PCI)或α1抗胰蛋白酶缓慢抑制。
PC被报道后不久,Discipio,一个PhD学生在位于西雅图(Seattle)的Davie实验室,发现另一种维生素K依赖性蛋白,被命名为蛋白S(PS)。几年后,Walker报道PS的功能是作为APC的辅因子。不久后发现此蛋白有两种血浆形式,游离的PS (30%~40%)和另一种与补体调节物- C4b结合蛋白(C4BP)结合成复合物的结合PS。血浆中C4BP的异构体由7个等同的α链和1个能结合PS的β链组成,每一个α链有一个C4b补体蛋白的结合位点。这些链以章鱼状形式深入各方。游离PS作为APC的辅因子参与下调凝血酶的生成反应。结合PS能和凋亡细胞表面的阴性磷脂相结合,使补体的活化局限在局部。APC对阴电荷磷脂膜表面的FⅧa和Ⅴa灭活极敏感。FⅧa(R336和R562)以及FⅤa(R306,R506和R679)是APC的敏感部位,被APC裂解成碎片使它们与各自酶(FⅨa和Ⅹa)的结合位点丢失。但是在Ⅹ酶(FⅨa-Ⅷa)和凝血酶原酶(FⅩa-Ⅴa)复合物中的FⅧa和FⅤa分别受各自的酶,FⅨa和FⅩa,部分地保护。
在阴性磷脂表面APC对Ⅹ酶和凝血酶原酶的调控需要PS的刺激。APC对Ⅹ酶复合物调控时的PS辅因子功能还需要受FⅤ的增效、协同作用。FⅧ血浆水平约为0. 7nM,较FⅤ的血浆水平20nM 低2个数量值顺序,而在凝血活化顺序中上游形成的Ⅹ酶复合物极少而下游形成的凝血酶原酶则是大量的。按此强大的逐级扩增的效应中,Ⅹ酶为关键酶。这可以解释PS的APC辅因子活性需要2个辅因子- PS和FⅤ来调控Ⅹ酶,而只需1个辅因子足以调控凝血酶原酶复合物。这亦提示FⅤ有促凝和抗凝的两种潜在表达。
蛋白C基因结构
PC只由肝细胞合成。人类PC mRNA含1800~1850bp核苷酸,<10%mRNA短于200个核苷酸。mRNA包括5'非翻译区(292核苷酸),PC前体物编码区(1383核苷酸),3'非编码区(292核苷酸)及多A尾上游19核苷酸处AATAAA信号或位于前一信号上游229核苷酸处ATTAAA信号。推测后一信号是决定少数较小类型mRNA。PC基因位于2号染色体q14- q21,全长11kb,有9个外显子,8个内含子。内含-外显交界按GT/AG规则分界。PC基因与FⅦ、FⅨ和FⅩ有35%~40%相似:四种因子蛋白的非催化部位都由不同外显子编码,拼接方式四种因子极相似,这提示它们有相同的蛋白进化起源。但四种因子基因的内含子序列及大小各不相同。外显子分别编码:5'非翻译区(外显子1),信号肽和前肽的6个氨基酸(外显子2),前肽其他部位包括γ羟基谷氨酸区(Gla区)(外显子3、4),两个EGF区(外显子5、6),轻链C端、活化肽和重链初始的27个氨基酸(外显子7),重链其他部分(外显子8、9)。
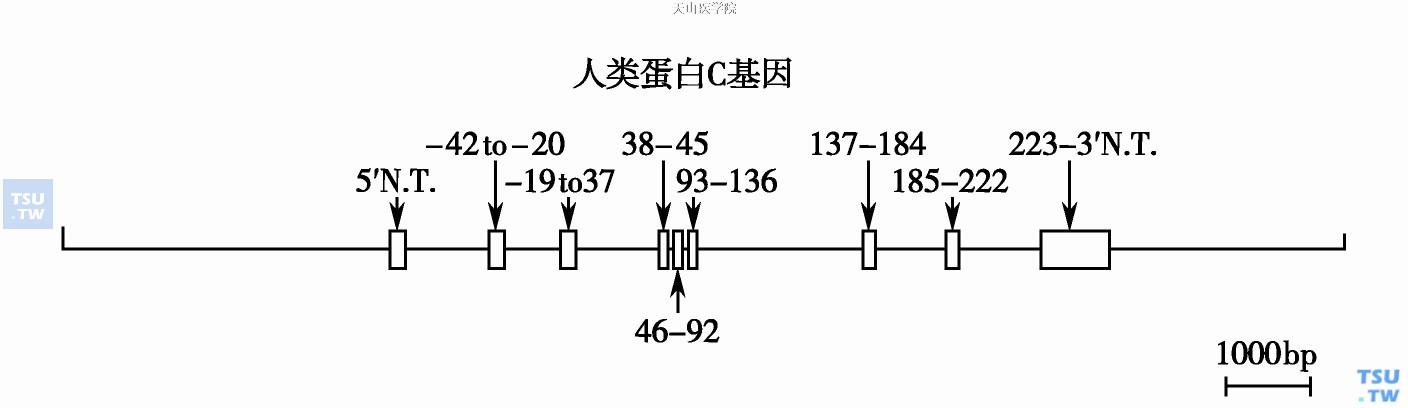
人类蛋白C基因;方框代表外显子部位及大小
蛋白C的分子结构及功能
前体蛋白
人类蛋白C在肝细胞线粒体合成,为糖蛋白。翻译后未经修饰的单链前体蛋白约Mr 62 000,含23%糖类。四个天门冬酰胺残基(1个在轻链,3个在重链)与糖基侧链相连。前体蛋白461个氨基酸长,含信号肽(33个氨基酸)、前肽(9个氨基酸)、轻链(155个氨基酸)、重链(262个氨基酸)、连接功能肽(Lys- Arg)(下图)。前肽是维生素K依赖性羧化酶识别部位。与其他维生素K依赖性凝血因子相似,在羟化酶参与下进行分泌前的γ羧基谷氨酸(Gla)加羧基作用,是蛋白翻译后的第一次修饰。分泌前被类β-羧肽酶(carboxypeptidase β- like enzyme)分别裂解Gly25- Thy24、Arg- 1-Ala1而除去信号肽和前肽;出线粒体之后又经高尔基体内Arg157- Thr158裂解,及血浆内Arg157-Lys156裂解而产生成熟型双链PC。PC血浆水平3~5μg/ml。约15%PC在血浆中仍为单链式。因PC酶原为单链式,故推理最初合成的PC在血浆中为单链,经裂解之后产生双链式PC。单、双链式PC活化后有相同的功能。人与牛PC有74%序列相同,核苷酸水平82%相同。
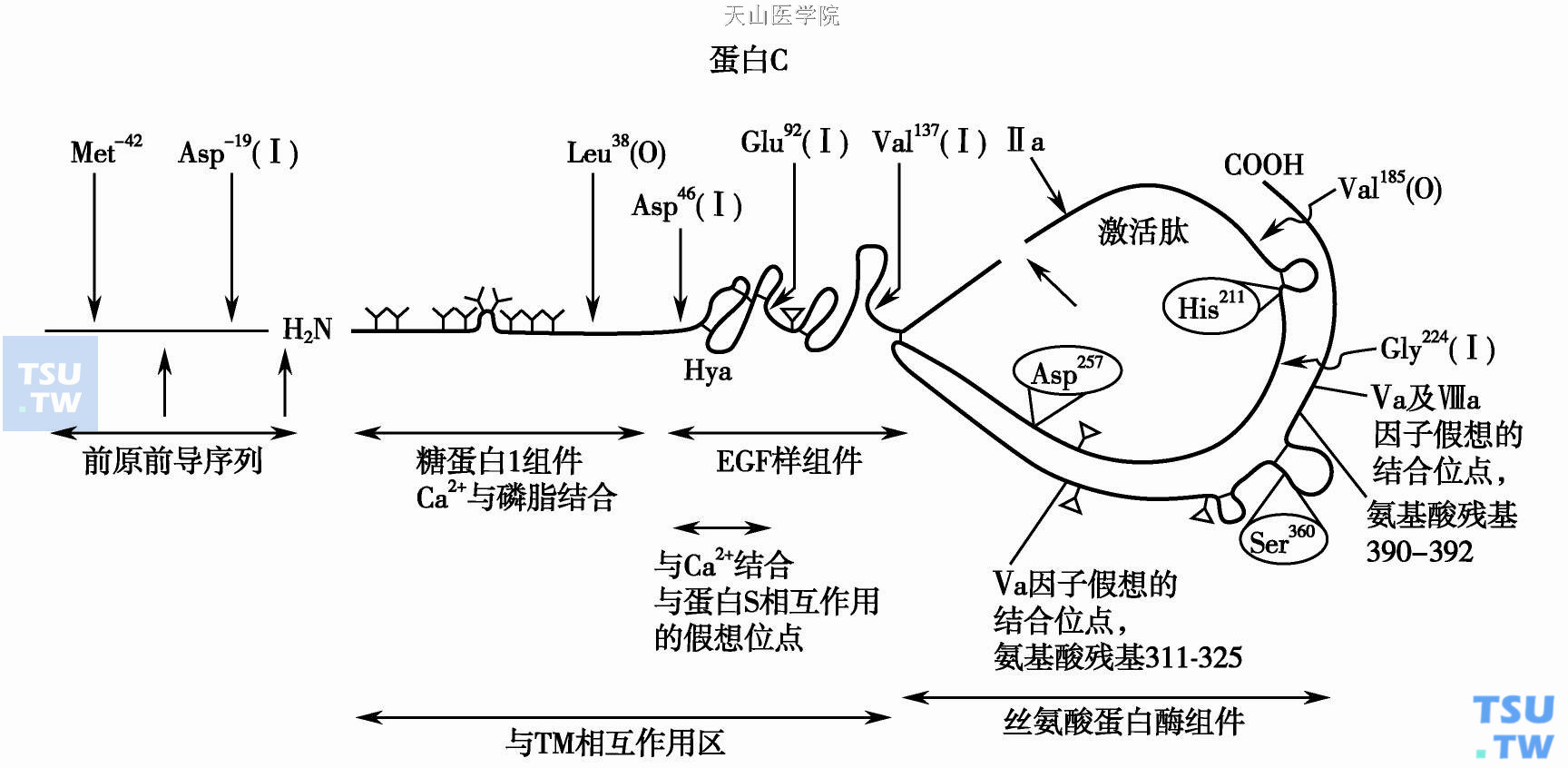
蛋白C的分子结构
蛋白C功能区
两种血浆PC分子名α和β,取决于N联结的糖基量。人体30%血浆PC重链Asn329为Asn- X- Cys序列,不接糖基,分子较小称β- PC;70%Asn329含Asn- X- Ser/Thy序列,接糖基,称α- PC。两种形式的血浆PC分子都可活化且具同样抗凝活性。
成熟PC分子轻链N端含Gla区,两个EGF区,重链含丝氨酸蛋白酶区。人类PC的Gla区含9个Gla残基,牛PC含11个Gla。该区确切的三维结构未明,但推测其功能与凝血酶原Gla区相似,能通过Ca2+桥联使PC分子云集在阴性电荷的细胞膜磷脂表面。在PC活化过程中Gla区能直接和TM相反应。6、7、19、20位Gla是APC活化所必需。Gla区中由6肽所组成的二硫键圈是APC抗凝功能所必需。第1 EGF区含8个半胱氨酸(Cys),其中Cys59、64在EGF区形成二硫键,是其他EGF区不具备的独特结构。第1 EGF区羟化所形成的赤藓(糖)-β羟化天门冬氨酸(Hya)是联结钙的独特部位。单个Ca2+与Hya结合可使PC构型发生改变,使它易被凝血酶- TM复合物所活化,并使之对凝血酶的单独作用不敏感。Hya是PC翻译后的第二种修饰。第1 EGF区能和PS的凝血酶区相反应,重链Asn97、202、313、329具有糖基化侧链,这些糖化位点是PC分泌所必需,30%Asn329不接糖基为β- PC。
蛋白C的激活及基础情况下活化蛋白C的功能
在无Ca2+的体外实验条件下,凝血酶、胰蛋白酶、蝰蛇毒等丝氨酸蛋白酶均可激活PC,但作用缓慢。上述丝氨酸蛋白酶中,凝血酶是唯一的PC生理活化物。PC在凝血酶作用下Arg12-Ⅱeβ链裂解使重链N端释出一个12肽,即使PC活化成APC。12肽称PC活化肽(PCP)Mr约1400,是PC活化的标志物,可用放射免疫或酶联法测定。内皮细胞表面结合着凝血酶调节蛋白(throbomodulin,TM)是凝血酶的受体。TM能与凝血酶结成1:1复合物而将经微循环灌注入的凝血酶和PC的活化反应加速2万倍。约有10万TM/内皮细胞,小血管表面比较大血管更能充分接触血流,故在基础情况下PC活化主要发生在微循环。微循环中APC能裂解血小板膜结合着的FⅤa;因为FⅤa是血小板膜上FⅩa受体,阻断FⅩa在血小板表面的积贮将进一步阻断凝血酶原的活化。APC并通过摧毁FⅧa生物活性而下调FⅨa活性,进一步限制FⅩa生成。因此,微循环中APC能在细胞表面通过调控FⅩa生成而有效阻断血小板、纤维蛋白沉积所产生的凝血活性,产生防栓抗栓功能。
PC活化过程取决于EGF区的钙结合部位。Ca2+在此部位的结合能使PC分子易被凝血酶- TM复合物所活化,并对凝血酶的单独活化不发生作用。APC抗凝作用需与内皮细胞或血小板表面结合着的PS相结合。APC的丝氨酸蛋白酶活化中心位于重链,与其他胰蛋白酶样蛋白酶的作用方式相似;对底物中的Arg残基有特异性,使存在于细胞膜表面的FⅤa、FⅧa发生蛋白溶解性降解。急性严重创伤引发的急性低灌注损伤,可使内皮受损。受损内皮表达过多TM,并能因此诱发急性创伤性凝血病。过度表达的TM使凝血酶从促纤维蛋白形成转向激活PC。这一后果将大大下调凝血酶的生成反应(低凝),并将促使原发性纤溶亢进和PC的耗竭。PC抗凝机制的耗竭最终将由低凝转向促使血栓形成(见第139章)。
血浆APC清除时间约15分钟。APC由肝脏直接清除,或被蛋白酶中和。正常血浆中各种蛋白酶抑制物,如PC抑制物(Mr 57 000,PCI又名PAI- 3)、α1抗胰蛋白酶、α2巨球蛋白、α2抗纤溶酶、PAI- 1都可中和APC活性。基础情况下,APC作为PC抗凝系统主要的酶具有生理性的抗血栓形成、抗感染和抗凋亡作用。
蛋白C激活的控制机制
几种不同的机制可以控制PC的激活。其一是由于凝血酶被抑制。血管壁的TM和类肝素协同来增强对凝血酶的抑制。当凝血酶和TM相结合,包括纤维蛋白生成和血小板活化在内的几种凝血反应被抑制。与此同时T- TM中结合的凝血酶被ATⅢ快速中和。T- TM复合物中的凝血酶较游离凝血酶与ATⅢ的中和反应更快速。加速机制依赖TM分子上存在的半乳糖氨基聚糖硫酸盐(又名chondroitin sulfate)。其二是由于PCI能快速抑制T 和TM的结合,而这种快速的抑制反应不需要半乳糖氨基聚糖硫酸盐的参与。T- TM中结合的凝血酶被快速灭活,以及PCI抑制T- TM复合物形成,这两种机制十分紧密地耦联能确保凝血瀑布反应中生成的凝血酶形成后不久,蛋白C的活化作用即被终止。TM和血管类肝素通过ATⅢ的一致行动来抑制凝血酶的相对重要性,在各种不同器官中有不同。例如脑微循环中TM只存在微量,几乎测不到。
凝血酶也能通过与TM结合的内存化而被清除。但是内存化的程度有不同,而且内存作用较凝血酶被抑制的速率为慢,使这一机制在调节中的作用不能确定。复合物中内存的T很快从TM解离,因此T和T- AT从循环清除都不以内存化清除为主要形式。
止血、感染中PC功能及APC抵抗
伤口的止血模型中,伤口部位过度生成的凝血酶,顺血流抵达内皮细胞表面结合着的TM,在Ca2+参与下将PC迅速活化成APC,复合物中的凝血酶本身失活,APC从复合物分离。然后APC在血小板或内皮表面下调凝血反应。未被激活的FⅤ和FⅧ对PC不敏感。炎症下,内皮表面TM减少使APC形成减少;细胞表面表达组织因子(TF)引起FⅩa增多,使血小板表面凝血酶生成增多。并且炎症下内皮-白细胞表达黏附分子(E或P选择素)致使白细胞参与局部加重血管损伤反应。炎症反应使血浆游离PS减少而进一步下调PC系的抗凝调控功能。因此炎症情况下诱生的损伤将促发血栓形成。狒狒的脓毒血症实验中先输注APC或PC能预防致死量(LD 100 doses)大肠杆菌的打击及DIC。但输注FⅩa抑制物,将激活部位封闭,仍能产生大肠杆菌的致死效应,提示APC、PS能在体内限制感染促使凝血系活化。
病理状态下,PC抗凝系的缺陷可引发高凝状态、血栓形成、缺血性卒中和高脂血症。PC缺乏症是一种少见的先天性易栓症,好发静脉血栓和习惯性流产,纯合子产生新生儿暴发性紫癜。先天性抗APC见于FⅤ- Leiden病。患者变异的FⅤ不能被APC裂解。获得性抗APC见于FⅧ血浆水平升高以及华法林抗凝治疗初期,反常的凝血活化能引发皮肤坏疽(即warfarin necrosis)。
蛋白S的基因
蛋白S基因是含有15个外显子的复杂基因。蛋白S缺乏病例的分子基础,至今只有少数能明确。这是因为PS存在假基因使分析复杂化。此外,PS体内的抗凝功能相当重要,但是在体外纯实验系统中只呈现2~3倍地增加抑制。再说,某些患者PS低水平是由于和C4b- BP结合的增加。Schmeidel等(1991)报道PS基因分析手段。国际血栓止血协会(ISTH)学术与标准化委员会血浆凝血抑制物分会(1997)总结报道了PS的基因缺陷的分子基础。
凝血酶调节蛋白的基因及T- TM复合物对纤溶功能的调节
TM是一种多功能蛋白,TM缺乏与血栓形成相联系。近年来的临床研究提示TM 1418位C/T两形性引起第6 EGF区丙455-缬(Ala455- Val)替代,可能和年轻人的心肌梗死有关。瑞典报道TM基因变异的C等位基因频率在存活的年轻心肌梗死患者中为5%,明显高于正常。这种方式的变异可在TM基因许多部位见到。TM缺乏内含子,因此TM基因变异的分析方法很简单。
Ireland等(1977)小宗研究报道TM基因5'调节区的变异在心肌梗死患者中为5%而对照组为1%。这一小宗研究表明TM启动子区多种变异的表达与动脉血栓形成相关。
近年来鉴定获得一种血浆羧肽酶B,称为凝血酶激活的纤溶抑制物(thrombin activatable fibrinolysis inhibitor,TAFI)。凝血酶能将此蛋白缓慢地激活,而TM则能加速这一过程。T- TM加速TAFI活化和TTM加速PC的活化有着相同的速率。TAFIa能裂解、除去羧基末端的精氨酸或赖氨酸残基。当这一过程发生在纤维蛋白凝血块中,能使纤维蛋白凝血块抗纤溶。
TAFIa也能影响其他一些介质的活性。如补体C5a(一种过敏毒素)羧基端精氨酸残基在活性上起重要作用。TAFI的调节功能尚未完全阐明。但是在炎症时输注TM能减少纤维蛋白沉着和炎症器官的损伤。这提示TM体内功效有显著的抗炎症和抗凝作用。创伤性急性低灌注损伤使内皮过度表达TM,将使凝血酶从促纤维蛋白形成转向促PC活化,从而产生纤溶亢进,最终因PC抗凝系衰竭而产生血栓形成。
内皮细胞蛋白C受体(EPCR)的基因
EPCR基因的可能变异与人类疾病间关系尚无确切信息。但是血浆可溶EPCR显著增高与血管受损性疾病如红斑狼疮的关系已明确。
总结:近年资料指出PC、PS和TM是凝血过程的重要抑制物。这一系统被炎症介质下调、减弱,被创伤性急性低灌注损伤加强至耗竭。PC、PS完全缺乏者有严重的血栓并发症。炎症介质使这一系统的能力下降,将不能使凝血酶转换功能,即从强的促凝酶转变为一个强的抗凝启动酶。
