
正常时,甲状旁腺素(PTH)由主细胞和嗜酸性细胞合成和分泌。完整的人PTH为一单链蛋白质,含84个氨基酸残基,分子量9500Da。循环血中的PTH具有显著的不均一性(heterogeneity),包括多种PTH肽段,如PTH1~84及其在主细胞、肝、肾等组织裂解而来的PTH的N端片段(PTH-N)和PTH的C端片段(PTH-C,6~7kD)。PTH-C虽无生物活性,但分子量较大,占PTH总量的4/5左右,其半衰期亦较PTH-N长。PTH中段片段(PTH-M)和C端PTH片段一样,从肾脏清除。此外,甲状旁腺还分泌少量的PTH原(pro-PTH)、前PTH原(prepro-PTH)和PTHrP。人prepro-PTH的分子结构见下图。
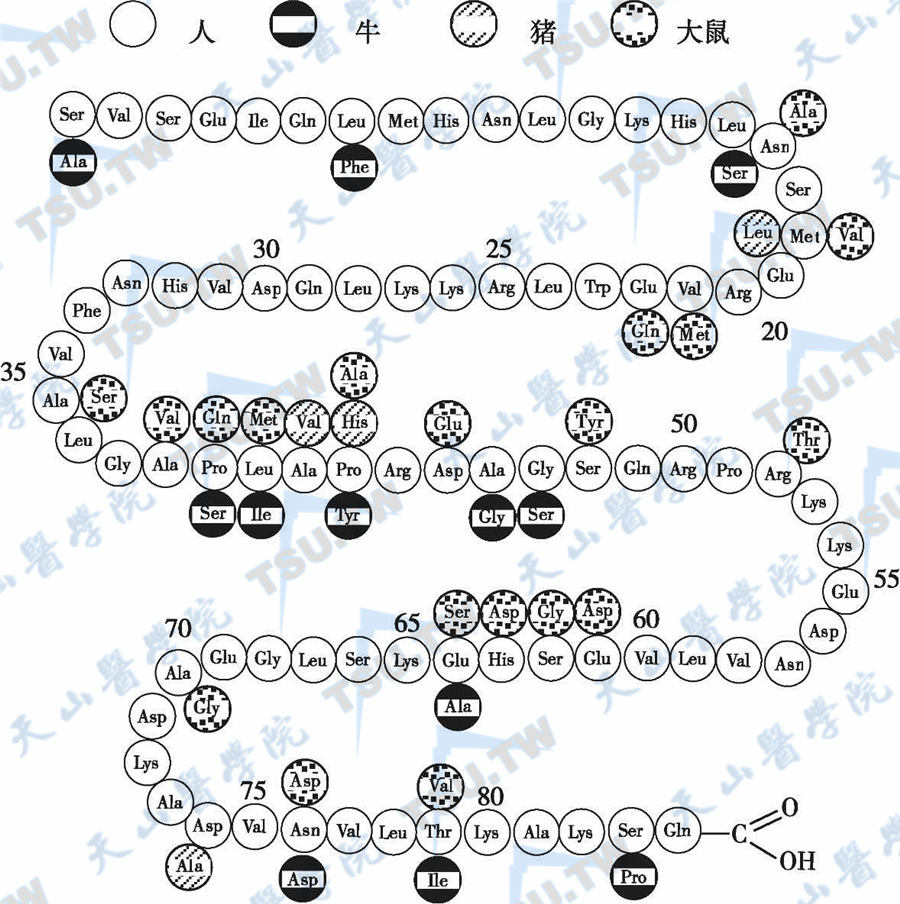
甲状旁腺素的分子结构
甲状旁腺素生物活性集中在1~34位肽链上
PTH的N端为PTH的活性端,第1~34个氨基酸残基片段(PTH-N1~34,2~3kD)的生物活性已达到PTH1~84所具有的全部活性。羧基端不具生物活性(下图)。
不同种属甲状旁腺素的生物活性(U/mg)比较
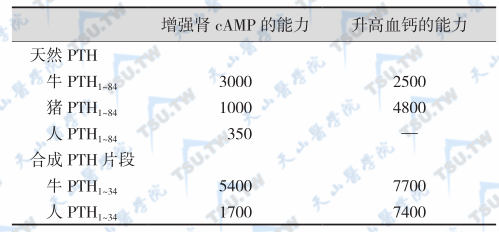
少于34或多于84个氨基酸序列的PTH片段也有一定的生物活性,但如果缺少第1位氨基酸,则活性明显减弱;如果缺少第1位及第2位氨基酸或少于27个氨基酸,则活性几乎完全丧失。PTH能刺激cAMP活性升高的最短片段为PTH1~27,并且主要与残基1和2有关。在氨基端的1~34个氨基酸序列中,人与牛有3个氨基酸不同,与猪有1个不同,人与鼠有4个不同;人与其他物种PTH1~84氨基酸序列中,人与牛有12个氨基酸不同,与鼠有19个氨基酸不同。人与猪仅有4个氨基酸不同,人与猪的PTH1~34保守区完全一致。由于氨基酸的组成不完全相同,多次给人注射牛或猪PTH后,会产生抗PTH抗体。
甲状旁腺素表达和分泌受许多因素的调节
人类PTH基因位于11号染色体短臂,包括3个外显子和2个内含子,并与降钙素基因邻近。限制性酶切分析人PTH基因,并用变性梯度凝胶电泳技术证实,不同个体的酶切片段呈多态性。这些遗传多态性对于遗传分析与PTH结构变异有关的甲状旁腺疾病诊断有重要意义。
PTH基因
编码人、牛、大鼠、猪、鸡和狗PTH的cDNA均已克隆。有关的基因组DNA(genomic DNA,gDNA)也从人、牛和大鼠中克隆。这些基因均有2个内含子和3个外显子。起始RNA转录了内含子和外显子的信息,之后,来自内含子的部分RNA序列被拼接,由成熟的PTH mRNA翻译前PTH原(prepro-PTH)。第1个内含子实际上把mRNA 5′端非转录区与基因的其他部分隔开;第2个内含子将编码“prepro”区域的部分与成熟PTH部分隔开。3个外显子也因此而分成不同的功能区。
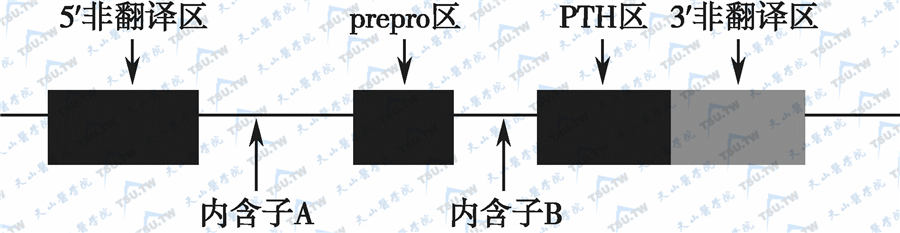
甲状旁腺素基因的结构示意图
注:外显子用长方形表示,A、B所示处为内含子区
第1个内含子在人类最大,约3400bp。第2个内含子在人、大鼠和牛约为100bp。哺乳类动物种属间的PTH基因结构有较高的同源性。人与牛约有85%相同,人与大鼠约有75%相同,而3′端非编码区却截然不同。人和牛有2个功能性TATA转录起始位点(大鼠只有1个)。位于PTH 84位谷氨酸编码后为终止密码子,故不会有羧基端延伸可能。PTH基因是典型的真核基因,具有始动RNA合成、RNA拼接、多腺苷化的全套序列。PTH基因在不同组织表达不同的原因可能还与基因不同位点胞嘧啶甲基化能力下降有关。有人发现,DNA在近PTH基因处的CPG序列甲基化减退。
PTH表达的调节
PTH基因受许多因素的调节。如维生素D(VD)、钙、磷、蛋白激酶A和C、性激素等。VD抑制PTH基因转录,使mRNA明显减少,PTH分泌减少,此与VD影响5′端侧翼区有关。另外,VD可使其在甲状旁腺的VD受体(VDR)基因表达增加,因而使VDR蛋白合成增加,VD 对PTH基因的作用被放大而产生显著作用。
现已合成了一些有生物活性但不升高血钙的VD类似物,如22-oxa-1,25-(OH)2D。这样可以减少PTH分泌,又不升高血钙。VD降低PTH基因转录的作用可治疗慢性肾病,预防继发性甲旁亢的发生。9-顺式视黄酸也有降低PTH mRNA的作用,与VD合用则有相加作用。新一代的VD类似物显示了更好的受体选择性,抑制PTH分泌的作用更强而促进肠钙磷吸收的作用更弱。西那卡塞(cinacalect)为拟钙类似物(calcimimetics),可增加透析患者的骨密度而不升高血钙。终末期肾病患者补充VD除治疗继发性甲旁亢,还有多种益处。除补充活性VD外,可考虑使用VD受体激活剂帕立骨化醇(paricalcitol)和25-(OH)D。如果血清PTH≥300pg/ ml,开始应用VD受体激活剂;如慢性肾病患者的血清PTH>70pg/ml,建议首先使用VD,恢复血25-(OH)D水平后再使用VD受体激活剂。VD受体激活剂,尤其是选择性VD受体激活剂(selective VD receptor activator)可抑制炎症、血栓栓塞和血管平滑肌细胞增殖与钙化。并可降调节肾素-血管紧张素-醛固酮(RAA)系统。
高血钙时抑制、低钙时兴奋PTHmRNA的表达;但没有证据表明血钙对降钙素基因也有调节作用。高钙血症对PTH的影响不如低钙血症。有关细胞外钙变化导致PTH改变的机制尚不明了。研究表明,Ca2+对PTHmRNA的调节主要在转录后水平,并且与钙受体和3′端UTR(untranslated region,UTR)结合有关。磷可通过影响VD、钙或直接影响PTH的基因表达。高磷血症直接刺激PTH分泌并使PTHmRNA升高。雌激素和孕激素对PTH基因表达均有调节作用。去卵巢鼠使用雌二醇后,PTH基因表达明显升高,但血钙和VD水平没有变化。蛋白激酶A和C对PTH mRNA也有调节作用。而且有证据表明蛋白激酶C对于PTH基因的正常表达是必需的。
甲状旁腺素合成与代谢差异导致分子不均一
血液循环中70%~95%的PTH是没有活性的C端PTH片段,5%~30%为PTH1~84。N端PTH片段可能是肝脏水解PTH1~84的产物,具有与PTH1~84一样的生物活性。肝脏产生的N端PTH片段迅速在原位降解,如果N端PTH片段在活体内要承担生物功能,必须提高自己在靶位点的局部产量,但目前尚缺乏可信的证据。
PTH合成和分泌
PTH基因经转录后,由胞质核糖体合成115肽的前PTH原(prepro-PTH),随即在粗面内质网池间隙经特异性肽酶裂解。前PTH原的“信号肽”序列位于N端。此25个氨基酸的疏水前导肽(“前”信号序列)裂解后,该序列即与粗面内质网上的“信号识别区”结合,并很快进入粗面内质网腔内,在穿过内质网膜时,该信号序列被迅速灭活。同时prepro-PTH即分裂成90肽的PTH原(pro-PTH)。随后,pro-PTH在高尔基复合体经酪蛋白酶及羧肽酶裂解进一步裂解为分泌型PTH。即由氨基端切下6个碱基(“原”序列),成为含84个氨基酸残基的成熟激素(分泌型PTH)。后者是激素分泌的主要形式,贮存在胞质内致密的分泌颗粒或微泡中。据估计,其储存量以最大速度分泌可持续1.5小时。在机体需要时,经胞吐释放入血。部分PTH在胞质内可降解成各种活性低或无活性的小分子片段,以调节机体对活性PTH的需要。
PTH分子形式
早期放射免疫测定法证实血液循环中的PTH与甲状旁腺中的不一样。PTH1~84和C端片段是主要成分。完整的PTH1~84在肝脏(占70%)及肾脏(占20%)分解成为有生物活性的N端片段(PTH1~34)、无生物活性的C端片段及中间片段。可用于PTH测定分析的有4种:全分子PTH(PTHl~84)、PTH-N(PTHl~34)、PTH-C和中间段PTH(PTH-M),其中PTHl~84和部分PTH-C是腺体直接分泌的,其余片段(包括部分PTH-C)都是PTHl~84在肝脏的裂解产物。各种分子形式的特点见下表。
血浆PTH的不均一性
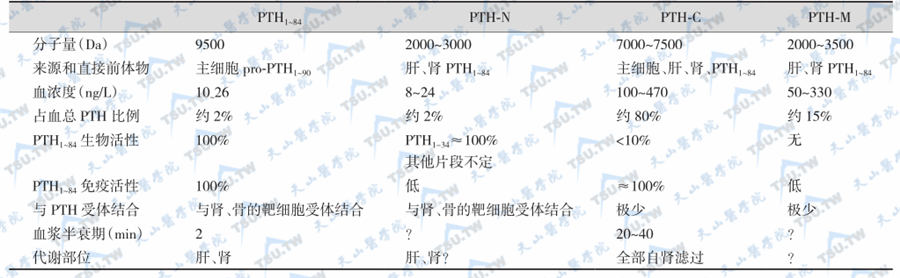
血液循环中可能还有从甲状旁腺细胞释放出来的prepro-PTH、pro-PTH及其他片段。这些片段绝大多数缺乏生物活性,pro-PTH的生物活性只有PTH1~84的2%~3%。但这些前体或片段可参与抗PTH抗体的免疫反应。血浆中PTH的多种形式构成了血PTH的不均一性。肾癌、肺癌及肝癌等均可分泌PTH或pro-PTH(异位PTH分泌)。
PTH代谢
PTH主要肝肾代谢,主细胞内的PTH代谢为血钙的稳定提供了另一条途径。循环血PTH的不均一性不但与PTH的外周代谢有关,也与甲状旁腺本身分泌的PTH片段可变性有关。甲状旁腺内PTH的降解是钙调节PTH分泌量和种类的一种机制。低钙血症时,PTH降解率下降,故完整PTH的分泌量增加;高钙血症时,PTH降解增加,完整PTH的分泌量减少。不过,此时PTH-M和PTH-C与PTH1~84的比率增加。PTH-C在高钙血症时的含量增加并不是由于甲旁亢所致。
前PTH原信号序列突变导致甲状旁腺功能减退症,这说明该信号序列对于维持PTH作用十分重要。6个碱基(“原”序列)的作用尚未明了,可能与信号序列发挥正常作用以及“前”信号序列的正确裂解有关。含84个氨基酸的成熟激素将在分泌囊泡和颗粒中浓缩。有关颗粒亚结构的形态学资料表明,颗粒中既含有甲状旁腺激素也含有蛋白酶(羧基化酶B和H,carboxylase B and H)。这种现象可解释为什么从甲状旁腺分泌的PTH有些有C端,但没有N端。由于C 端PTH对钙内环境的调节作用不明显,因而反映了体内的灭活途径,是PTH合成、分泌和生物作用的重要的调节机制。高钙血症时,PTH分泌减少而且分泌的多为无活性的C端PTH。
PTH在肝脏的清除较复杂,Kupffer细胞可以非饱和性摄取PTH,分解成小分子片段且容量较大,在33、34、36及37位切断PTH1~84,形成C端PTH片段。PTH半衰期约2分钟。PTH的这种快速外周代谢不受血钙或1,25-(OH)2D的调节,分泌的PTH仅约1%到达靶器官。肾脏是清除C端PTH片段的主要部位,肾小球滤过率下降时该片段在体内堆积。即使肾功能正常,该片段的半衰期也比PTH1~84要长5~10倍。测得的血PTH浓度取决于PTH的分泌率和血液对迅速变化的激素分泌率反应的快慢。
钙依赖性甲状旁腺素分泌是低钙血症的有效防御机制
血Ca2+对PTH的调节
PTH调节血清钙磷水平和骨代谢。甲状旁腺细胞促发PTH分泌的因素是细胞外Ca2+降低而非升高,这一点与其他任何激素的分泌模式是不同的,这是因为有钙受体(calcium receptor,CaR)的参与。甲状旁腺细胞的另一个显著特点是PTH分泌颗粒所含的PTH量很低,因此,其分泌量的调节主要依赖于瞬时的PTH基因表达与甲状旁腺细胞增殖。
甲状旁腺细胞对Ca2+的反应与其他组织的细胞不同。低血钙可以兴奋甲状旁腺,而血浆离子钙浓度升高时则抑制PTH的分泌。其量效曲线呈S形(曲线中点处的钙水平称为调定点)。正常情况下,PTH分泌以调定点方式控制来维持血清离子钙在一个很窄的范围内。低于调定点时刺激PTH分泌,高于调定点时抑制PTH分泌。但是,PTH的分泌速度有一定限度,血清钙为7mg/dl时,兴奋作用最大,血清钙为10.5mg/dl时,抑制作用最大,高于或低于此水平不产生更大的作用。钙对激素分泌的作用发生很快(数分钟内),低钙对于PTH合成的刺激较慢。高钙浓度时,可使细胞内合成的PTH降解,而且可能释放无活性片段。短时间内,细胞外钙主要调节PTH的分泌而不是其合成;但数小时到数天后,细胞外钙增加可抑制PTH基因转录,反之亦然。细胞外钙减少时促进PTH基因转录,长期的低钙血症刺激甲状旁腺细胞增殖、肥大。通常情况下,当血液中离子钙浓度增加时,PTH的分泌下降。其量-量反应曲线呈S形(图A),决定该曲线的参数主要有4个(图B):最大分泌率(图B中的A线)、曲线中点的斜率(B线)、中点钙浓度(C线)和最小分泌率(D线)。
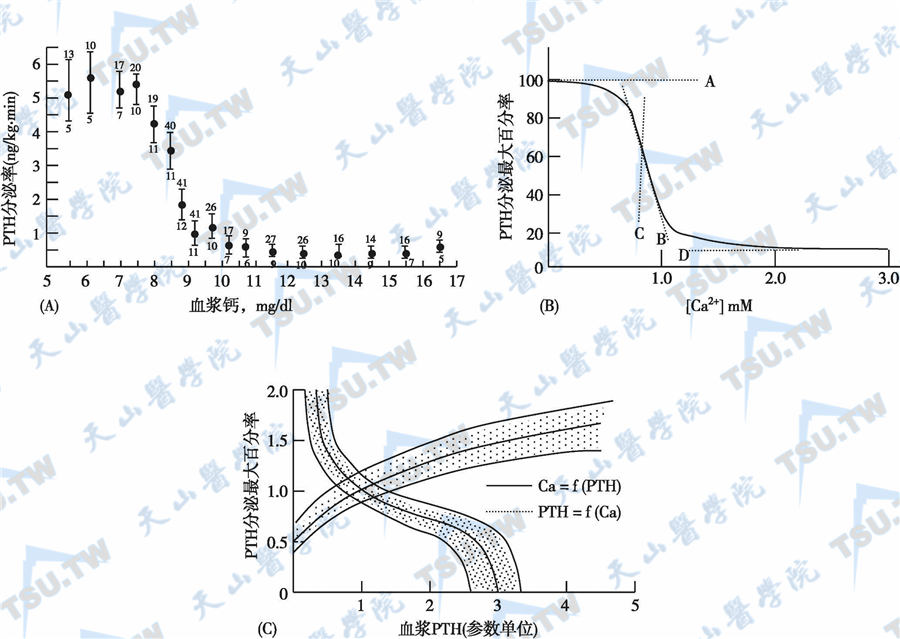
甲状旁腺素(PTH)与血钙的变化关系
甲状旁腺细胞对血钙的反应性决定了曲线呈S形,但并不是唯一决定每一个维持生理稳态的具体调定点。该点一般位于曲线中点和底部之间的区间,并可通过检测靶器官对PTH的反应而测出(图3-14-10C中的实线)。稳态(steady state)是图3-14-10C中PTH和钙曲线的交叉部分。S形曲线代表了甲状旁腺的几个生理特性。例如,最小分泌率很低但不等于零,最大分泌率反映甲状旁腺对低钙的储备能力。由于正常人的稳态位于曲线中部和底部之间的区域,故对低钙血症的反应比对高钙血症的反应要显著得多。研究表明,甲状旁腺细胞对血钙的绝对值和血钙的变化速度均有反应。快速下降的血钙比缓慢下降所造成的刺激要强。这样,为机体提供了另一防止低钙血症的保护机制。
钙受体对PTH的调节
钙受体(CaR)是G蛋白耦联受体家族中的成员。分布在甲状旁腺细胞的多种细胞表面。它可以兴奋磷脂酶C并阻断cAMP产物的刺激作用。该基因的失活性突变可导致家族性低尿钙性高钙血症的发生。该基因单拷贝转基因鼠的甲状旁腺钙受体有缺陷。能激活钙受体克隆的拟钙(calcimimetic)复合物可抑制PTH分泌,并有望成为原发性和继发性甲旁亢的治疗药物。大多数细胞均不受细胞外钙浓度波动的影响,保持着非常低的细胞内钙水平。但甲状旁腺细胞的细胞内钙水平却在一定范围内随细胞外钙浓度的波动而变化。钙活化细胞表面的钙受体使细胞内储存的钙释出,同时细胞膜上的钙通道开放,使细胞内钙水平升高,后者通过目前尚不清楚的调节机制使PTH分泌减少。
血磷对PTH的调节
PTH降低肾脏对磷的重吸收,调节血磷浓度;反过来,肾病时血磷升高也影响甲状旁腺,导致继发性甲旁亢。该调节独立于磷对钙及1,25-(OH)2D的作用,磷直接调节1,25-(OH)2D的生成,磷与钙形成复合物而影响血钙,间接对甲状旁腺起调节作用。低磷血症直接使PTH 表达下降,而高磷血症则相反。低磷血症还抑制甲状旁腺细胞增殖,但不清楚磷是否直接影响PTH的合成和分泌。快速给予磷制剂刺激PTH分泌,这一作用是继发于Ca2+下降所致。高磷血症抑制VD活化,促使Ca2+进入线粒体,降低细胞外Ca2+水平。肾功能不良时,钙磷代谢紊乱,VD不能在肾脏进行1α位羟化,引起低钙血症,刺激甲状旁腺增生。
昼夜节律对PTH的影响
生理情况下,PTH的分泌有昼夜节律性,白天的血PTH浓度平稳,夜间20点及凌晨4点出现两个宽的高峰,其中后1个高峰值可持续到上午8~10点才降至基础水平。所以,临床上鉴别正常人和轻度甲旁亢应在上午10点后抽血测定PTH。正常人PTH分泌呈昼夜节律的机制尚不清楚,原发性甲旁亢患者此节律消失。
特殊情况下的PTH分泌
新生儿由于胎儿血钙浓度高于母体,其PTH分泌处于抑制状态,出生后血钙呈生理性下降,PTH分泌逐渐上升。女性的PTH分泌随月经周期而轻微波动,排卵期有一PTH浓度高峰,其峰值比卵泡早期和黄体后期高30%~50%。孕晚期(24周以后)血PTH浓度逐渐上升,以动员母体骨钙供给胎儿;虽然哺乳期每日经乳汁泌钙达1g,但其甲状旁腺分泌功能无明显变化,其机制不明。
其他因素对PTH的调节
- 血清镁:细胞外液镁离子浓度升高也像高钙浓度一样可抑制PTH分泌。但是低血镁不同于低血钙,它可抑制PTH的分泌和作用。镁的调节能力远不如钙,只有在重度高镁血症、低镁血症时才可引起PTH分泌受影响。严重镁缺乏不但减少PTH的释放,而且使PTH与受体结合力减弱。
- 锶盐与锂盐:锶盐(strontium)抑制PTH的释放,而锂盐(lithium)兴奋PTH的分泌,锂盐还降低细胞外钙抑制PTH分泌的敏感性。
- 维生素D:如维生素D (VD)在体内不能转变成足量的活性分子,可发生肠钙吸收不良,导致低钙血症;另一方面,PTH没有VD的“允许”作用,不能使靶器官产生相应量的cAMP,因此不能发挥PTH的生理效应。所以VD缺乏或不能活化是继发性甲旁亢的重要病因。VD的活性代谢产物1,25-(OH)2D抑制PTH合成和分泌。肾衰竭时,1,25-(OH)2D降低是引起PTH增高的重要原因。1,25-(OH)2D抑制PTH基因转录,此乃PTH促进VD活化的负反馈机制。
- 降钙素:降钙素抑制骨细胞及破骨细胞的骨吸收作用,增加成骨细胞活性而使钙磷沉积于骨骼,血钙和磷降低。降钙素对肾小管的作用与PTH相似,使肾小管重吸收磷减少,尿磷排出增多,但降钙素也减少肾小管对钙、钠、钾及镁的重吸收,抑制肠钙吸收,降低血钙,并通过降低血钙及阻止PTH与受体的结合,刺激PTH分泌。
- 排磷素:排磷素(phosphatonin,FGF23)与PTH的关系仍未明了。原发性甲旁亢患者的血磷降低,血klotho和FGF23升高,而此时PTH可能正常或仅轻度升高。由此提示,排磷素对PTH有反馈抑制作用。
- 其他因素:雌激素降低骨基质分解、抑制骨对PTH的应答作用。大量皮质醇抑制肠钙吸收,抑制肾小管重吸收钙与磷,造成低钙血症。皮质醇直接刺激PTH分泌,造成低磷酸盐血症,骨吸收加快。肾上腺素、β肾上腺素能兴奋剂、多巴胺、胰泌素、泌乳素及生长激素等使PTH分泌增多,腺体增生。普萘洛尔(心得安)抑制PTH的分泌。
