
肾上腺作为体内重要的内分泌腺体,由结构和功能各异而又相互联系的皮质和髓质组成。皮质由外至内分为球状带、束状带和网状带。球状带合成和分泌醛固酮,属于肾素-血管紧张素-醛固酮(renin-angiotensin-aldosterone,RAA)系统,主要参与血压和体内水盐代谢的调节;束状带主要合成和分泌皮质醇,属于CRH-ACTH-皮质醇轴(HPA轴),主要参与应激反应、物质代谢和免疫功能的调节;在通常情况下,网状带主要合成和分泌类固醇性激素(如雄烯二酮、去氢异雄酮、孕酮和雌二醇等),主要参与性腺(睾丸或卵巢)功能和代谢的调节。肾上腺髓质主要由嗜铬细胞和神经突触组成。髓质相当于交感神经的节后部分,是APUD细胞的集聚体,除合成和分泌儿茶酚胺(catecholamine,CA)外,还可分泌许多肽类与胺类激素,以旁分泌/自分泌途径调节局部的各种功能。下面先说说下丘脑-垂体-肾上腺轴:
CRH刺激垂体ACTH和生长抑素分泌
CRH神经元
中枢神经分泌CRH的细胞主要分布于室旁核和杏仁核,其轴突多投射到正中隆突处或下丘脑的基底面,但CRH神经纤维和CRH阳性物可分布于海马、中脑、松果体等许多区域。HPA轴的主要生理功能是调节机体对各种应激的反应,当机体处于应激状态时,中枢神经CRH神经元的CRH和AVP (ADH)表达增多。
CRH分泌
CRH的分泌受中枢交感神经的调节。交感神经兴奋时,CRH分泌增多。目前,关于CRH分泌的调节机制尚未完全明了,概括起来有下列特点:
- 应激状态下的ACTH分泌主要受CRH和AVP的兴奋,CRH和AVP在接受应激信号后,可单独或由CRH/AVP神经元共同表达CRH和AVP,心房利钠肽(ANP)和脑钠肽(CNP)对HPA轴有抑制作用,内源性致焦虑因子为ANP受体的抑制剂,因而AVP与ANP对CRH的作用是互相拮抗的。
- 褪黑素(melatonin)减弱肾上腺皮质对应激的反应性,减少垂体ACTH分泌;另一方面,CRH又抑制松果体褪黑素分泌,因此,CRH和褪黑素又组成负反馈调节系统。
- 除兴奋HPA轴外,CRH还影响行为,兴奋交感神经系统和抑制食欲。
- 胎盘CRH是启动分娩的刺激物。
- IL-1和IL-6也可调节CRH的分泌,而瘦素可调节CRH、GHRH、生长抑素、TRH等的基因表达,是能量代谢的主要中枢性调节因子。
- CRH受体-1拮抗剂(如antalarmin)可抑制HPA轴、交感神经和交感-肾上腺活动,可用于治疗因CRH分泌过多所致的精神病、心血管疾病或生殖系统疾病。
- CRH结合蛋白(CRH-BP)除具有一般激素结合蛋白的功能外,还能直接调节CRH和ACTH的翻译、分泌和生物学作用,并与CRH-CRH受体一起组成局部调节网络,调节HPA轴功能和炎症反应。
- 下丘脑和垂体存在一组肽类或胺类活性物质,它们具有拮抗CRH,抑制ACTH合成和分泌的作用。目前认为,生长抑素、多巴胺、ANP和前TRH原(prepro-TRH)均可作用于ACTH细胞,抑制POMC基因表达,被统称为促肾上腺皮质素释放抑制因子(CRIF)。
- 腺垂体也含有大量的神经纤维,某些神经纤维与激素分泌细胞有突触联系,使神经系统可直接影响垂体前叶的激素(主要是ACTH)分泌。
CRH对ACTH和生长抑素的调节
经垂体-门脉系统或一些目前尚未完全阐明的途径作用于垂体ACTH细胞,刺激ACTH分泌。如下丘脑的室旁核或正中隆突受损,应激动物的ACTH释放明显减少。CRH还可促进局部生长抑素分泌,抑制LH的释放。给人或动物静脉注射CRH后,血浆ACTH、β-内啡肽明显升高,因此可用CRH兴奋试验来了解垂体的ACTH贮备功能,并对CS的病因有鉴别意义(本章第114节和第115节)。
皮质醇反馈抑制CRH和ACTH分泌
皮质醇对垂体ACTH的抑制作用与对下丘脑CRH的抑制作用均较明显,其中可能以抑制ACTH的作用为主。皮质醇抑制ACTH的作用可分为延迟反馈抑制和快速反馈抑制两种类型。在输注外源性皮质醇后数分钟内可将ACTH的分泌抑制80%,但这种快速抑制作用约于20分钟后消失;数小时后又再次出现对ACTH的反馈抑制作用。快速抑制反应的机制未明,可能与糖皮质激素的膜结合活性及其引发的ACTH分泌抑制有关。延迟抑制反应是糖皮质激素与核受体作用,激活相关基因表达,mRNA转录及新的蛋白质合成的结果。糖皮质激素可诱导脂调素(lipocortin,LC)合成,LC是磷脂酶A2的抑制物,可抑制POMC基因转录和ACTH合成。
糖皮质激素的作用十分广泛,几乎影响到体内所有的代谢过程和所有的组织功能,但不同生物学作用的途径是相同的,即均通过膜联蛋白(annexin)发挥作用,因而可将其视为糖皮质激素的第二信使。腺垂体的ACTH细胞、GH细胞、PRL细胞、TSH细胞和LH/FSH细胞均可表达高亲和力annexin受体,因而糖皮质激素对这些垂体激素的合成和分泌均有调节作用。但也有人认为,只有垂体的滤泡星形细胞(folliculostellate cells)可合成和分泌annexin-1,并通过旁分泌机制调节垂体激素分泌细胞的活动。在糖皮质激素的作用下,靶细胞合成annexin-1,并从浆膜内侧向细胞膜外侧转位,这可能也是糖皮质激素非核受体作用的途径之一。糖皮质激素的抗炎作用、抗异体排斥反应和免疫抑制作用均与annexin有关。在下丘脑,糖皮质激素对CRH的抑制作用由糖皮质激素受体介导。Ⅰ型糖皮质激素的受体活性可调节CRH和AVP表达;而Ⅱ型糖皮质激素受体主要参与应激性ACTH分泌调节。
在急、慢性应激情况下及一些精神性疾病时,中枢神经系统的糖皮质激素受体功能异常,此时可用DXM-CRH试验来了解ACTH的反应性,抗抑郁药可抑制CRH及AVP合成,而一些躯体疾病或感染性疾病(如HIV感染)常伴有HPA轴功能紊乱。肾上腺分泌的各种皮质激素(如醛固酮和皮质醇)、激素前体物及代谢产物均对中枢神经系统有一定影响,甚至可干扰脑电活动。而神经节细胞可合成和分泌孕酮及去氢异雄酮,并以旁分泌方式调节行为活动和中枢神经系统的学习、记忆与睡眠等生理过程。
ACTH和交感神经促进皮质醇分泌
皮质醇的分泌可分为“基础分泌”与应激状态下的“增量分泌”(应激分泌)两种情况。基础分泌是指机体日常活动时的一般性皮质醇分泌,而应激性分泌是机体根据应激的需要而增加的皮质醇分泌。肾上腺皮质醇的分泌调节具有以下特点:①ACTH促进皮质醇的分泌具有可饱和性,在达到最大兴奋值前,ACTH与皮质醇浓度有线性关系,但超过最大兴奋值,ACTH增加不再有更多的皮质醇分泌。欲达到肾上腺皮质最大兴奋反应所需的ACTH值尚未确定。根据大量的临床试验观察,一般使用1~5μg ACTH1~24即可达到对肾上腺皮质的最大兴奋,以前使用25U的ACTH1~39兴奋试验显得用量过大。②即使摘除垂体,肾上腺的皮质醇分泌并不完全停止,这说明肾上腺的皮质醇分泌尚具有非ACTH依赖性途径。现发现,交感神经可直接促进皮质醇的合成与分泌。此外,在肾上腺局部还有皮质醇分泌的旁分泌调节途径(见后述)。③应激状态下,可通过交感神经、交感肾上腺髓质-肾上腺皮质及CRH/ACTH途径促进皮质醇的合成和分泌,使血浆皮质醇在很短时间内显著高于基础状态。
ACTH的作用广泛,现已发现5种黑色素皮质素受体(MCR)。MCR属于G蛋白耦联受体家族中的成员,MC2R即ACTH受体,MC2R与ACTH结合后,激活腺苷环化酶,细胞内cAMP浓度升高,产生相应的生物效应。MC2R只在肾上腺皮质中表达,在束状带、球状带均广泛分布有MC2R,而网状带和肾上腺髓质的MC2R表达很少。IGF-1促进肾上腺MC2R的表达。IGF-1、IGF-2、EGF、FGF和TGF还可调节皮质激素的合成、分泌和肾上腺的发育与细胞凋亡。
促进肾上腺皮质细胞增生和活性
切除垂体的动物肾上腺皮质萎缩,补充外源性ACTH后,肾上腺皮质细胞增生肥大(以束状带和网状带增生为主),束状带的明亮细胞变为致密细胞,胞内脂质减少,线粒体肥大,一些与糖皮质激素合成有关的酶类以及碱性磷酸酶和酸性磷酸酶等的合成均增多,同时肾上腺血流灌注增加。
促进肾上腺皮质激素合成和分泌
ACTH促进皮质醇的合成和分泌。给人静脉注射ACTH后数分钟内即可见血中的皮质醇升高。此外,ACTH亦促进醛固酮的分泌(但其作用较弱)和肾上腺雄激素与雌激素的合成和分泌。
对神经系统的作用
中枢神经系统广泛分布有ACTH结合位点,结合位点密度以大脑皮质和下丘脑最高,其次为小脑、中脑、脑干和脊髓等处。ACTH对中枢神经的作用广泛,参与学习、记忆、行为、体温、心血管功能、神经修复、应激、止痛、镇静等重要生理活动的调节过程。例如,ACTH可拮抗阿片受体的作用,ACTH对正常动物行为的影响很明显,可降低情绪反应,诱发牵张反应(stretching reaction)等。ACTH亦促进周围神经的再生和修复,同时对生长发育中的中枢神经有营养支持作用。
对免疫系统的作用
ACTH是一种免疫调节激素。现证明,淋巴细胞和巨噬细胞可合成和分泌少量的ACTH,细胞合成的POMC虽然缺乏转录信号肽,但分泌的ACTH却可刺激肾上腺皮质细胞合成皮质醇,而且淋巴细胞还含有ACTH受体。看来,免疫细胞分泌的ACTH既可调节免疫细胞的功能,又可调节其他细胞的功能,皮质醇和DXM可抑制淋巴细胞的ACTH合成,这说明在免疫细胞中存在一套不依赖于垂体来源的ACTH调节系统。ACTH抑制T淋巴细胞依赖性抗原和非T淋巴细胞依赖性抗原引发的免疫反应,抑制T淋巴细胞γ-干扰素的合成及干扰素诱导的巨噬细胞活性。
旁分泌功能
CRH是一种主要的应激激素,在局部组织中CRH调节生殖系统的各种功能。例如卵巢和子宫内膜的CRH作为旁分泌和自分泌因子调节类固醇性激素的合成、分泌、排卵和黄体溶解等过程,胎盘“CRH钟”(CRH placental clock)决定了孕期和分娩的时间。
ACTH/皮质醇昼夜节律起源于下丘脑
ACTH和皮质醇的昼夜节律分泌来源于下丘脑的CRH周期性分泌,后者主要与生物钟和黑暗-光照(睡眠-醒觉)的周期性变化有关。
生物钟位于下丘脑前部的视交叉上核(suprachiasmatic nucleus,SCN),个体的发育、生长、环境变化和神经-内分泌“生物钟”现象都与SCN的活动有关,并受脑内神经递质,如5-羟色氨(hydroxytryptophan,5-HT)、胆碱能神经递质、兴奋性氨基酸、神经肽Y、脑啡肽、褪黑素、精氨酸血管加压素(AVP)及睡眠-觉醒肽(hypocretins)的影响,其中褪黑素在调节昼夜节律基因表达中起关键作用,而褪黑素分泌又是由光照和血清素能神经活动引起的。
血皮质醇的昼夜节律是垂体ACTH分泌节律性(来源于CRH分泌的节律性)的反映,与下丘脑和垂体的其他激素相比,ACTH和皮质醇的昼夜血浓度变化依赖于睡眠的程度低,暂时改变睡眠习惯对ACTH和皮质醇的浓度变化影响较小或无影响,但是ACTH和皮质醇的昼夜浓度差最为明显。一般糖皮质激素对垂体ACTH的反馈抑制以夜间最强,晨间最弱,如正常人在早晨服用DXM,其对内源性糖皮质激素的分泌抑制作用最弱,抑制持续时间也最短。相反,如正常人在晚上服用糖皮质激素,其对ACTH的分泌抑制作用最强,而且持续的时间也较长。同理,肾上腺皮质对美替拉酮(甲吡酮)的敏感性也以晚间为强,凌晨时较弱。这是多年来倡导隔日糖皮质激素治疗的理论基础,如1日数次等量投予外源性糖皮质激素,极易造成对肾上腺皮质功能的抑制。
HPA轴抑制因子维持内环境稳定
HPA轴抑制因子
体内存在许多可抑制下丘脑CRH或垂体ACTH合成与分泌的体液因子。在这些因子中,有的为旁分泌激素或细胞因子,有的则属于循环性内分泌激素;例如,抑制下丘脑CRH合成与分泌的因子有GABA、ACTH、β-内啡肽、CRH(自身调节)、瘦素(leptin)、P物质、生长抑素、催产素、NO、CO、脂调素、内皮素-1等;抑制垂体ACTH合成与分泌的因子有ANP、前TRH原(prepro-TRH178~199)、脂调素、肾上腺髓质素等。
为什么在体内已有糖皮质激素负反馈抑制CRH和ACTH情况下,还存在如此多的HPA轴抑制因子的生理意义未明。一种解释是机体随时都有应激性反应,这些抑制因子是HPA轴的一种保护性机制,由于应激的类型、方式和反应通路各不相同,为了达到精细调节的目的,生物进化中,逐渐形成了众多的拮抗或适应各种应激反应的调节系统,以维持内环境的稳定。
其他因素对HPA轴的影响
影响HPA轴功能的因素很多,其调节的综合目的是使机体适应内、外环境的变化。
- 妊娠:妊娠妇女的血ACTH升高(一般为正常高值),血皮质醇总量、游离皮质醇和CBG增高。分娩时可出现血ACTH和皮质醇的急剧升高。在妊娠期间,DXM不能满意抑制ACTH的分泌,这主要是孕妇血浆中的ACTH有一部分来源于胎盘(胎盘组织存在独立于母体的CRH/CRH-BP-ACTH-皮质醇/CBG系统)所致。
- 进食:进食后血ACTH升高(其机制未明),因此必须采取空腹的定时血浆标本测定血ACTH(受进食和昼夜节律的双重影响)。
- AVP和催产素:将AVP注入脑室内可引起ACTH分泌。动物实验表明,催产素也有类似作用,其机制未明。中枢神经组织存在ANP、脑钠肽(CNP)及其受体系统,可调节行为和ACTH的分泌。
- 中枢神经递质和神经调质:许多中枢神经递质或神经调质对ACTH的合成和分泌有调节作用。一般来说,引起CRH或ACTH分泌增多的因子有乙酰胆碱、5-羟色胺、吗啡类药物或内啡肽、AT-2、甘丙素、TRH、EGF、IL-1、IL-2、IL-6、TNF-α、LC、IFN-α和胸腺素等。血管活性肠肽(VIP)和垂体腺苷环化酶活化肽(PACAP)的氨基酸序列有高度同源性(其受体称为PACAP/AVP受体),广泛存在于神经纤维、下丘脑神经元及肾上腺组织(以髓质为主)中,垂体含有较高浓度的VIP。此两种旁分泌激素均刺激ACTH分泌(VIP主要增加CRH分泌,而PACAP直接促进ACTH释放)。VIP还与肾上腺皮质的ACTH受体(MC2R)亚型发生“串语”或与AT-2受体作用,促进皮质醇、醛固酮及儿茶酚胺的分泌(这些调节作用主要为病理性的,如炎症或应激)。CS的肾上腺皮质VIP增多,醛固酮增多症伴嗜铬细胞瘤的发病机制可能与VIP/PACAP基因的过度表达有关。抑制ACTH分泌的因子有去甲肾上腺素、γ-氨基丁酸(GABA,可能是DXM抑制ACTH分泌的介导物)、多巴胺、吗啡受体激动剂、P物质、TGF-β和NO等。
综上所述,HPA轴的调节途径和机制可表示如下图。
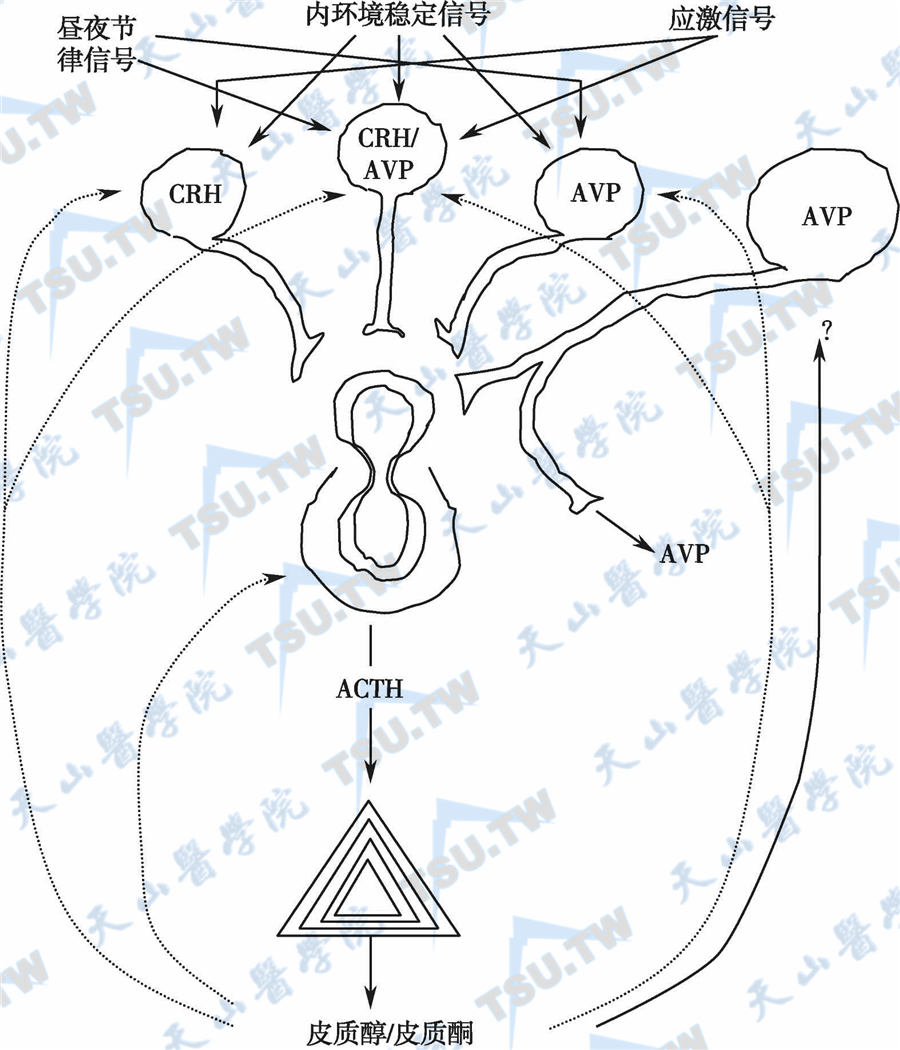
下丘脑-垂体-肾上腺轴及其调节
注: :表示兴奋;
:表示兴奋; :表示抑制
:表示抑制
肾上腺皮质和髓质参与应激反应并与神经调节密切配合
多种急性应激刺激可引起ACTH分泌的急剧增加,持续时间可达数小时之久。各种应激性刺激通过外周神经传入中枢神经系统,并将应激信号整合,汇集成神经递质性信号,作用于下丘脑的CRH神经元及其他相关神经元,CRH分泌增多,数分钟后血中的ACTH可升高4~6倍,同时伴血皮质醇浓度的同步性升高(8~10倍)。体液性应激性调节与神经调节密切配合,使机体适应应激反应的需要,提高机体的适应能力。但由于应激的性质、程度和机体反应能力的差异,应激刺激后个体的HPA轴变化可多种多样,除CRH外,参与急性应激调节的其他因素还有AVP、催产素、中枢性儿茶酚胺等。
肾上腺皮质和髓质均是调节应激反应的重要内分泌腺体。一方面皮质中含有嗜铬细胞,儿茶酚胺受皮质醇的调节;另一方面,肾上腺存在肾素和CRH的旁分泌调节机制,通过多种途径调节肾上腺皮质和髓质的应激功能。慢性应激时也有ACTH和皮质醇的调节参与。不过此时在各组织中,免疫因子的作用似乎比神经调节和HPA轴功能调节更为重要。慢性应激使肾上腺皮质的激素分泌具有耐受性和适应性(自身调节性),即使无ACTH的作用,糖皮质激素的分泌也持续增多。
