
载脂蛋白调节脂蛋白代谢
载脂蛋白是脂蛋白中蛋白质成分的总称,在脂蛋白的结构、功能与代谢等方面具有非常重要的作用。目前已发现的载脂蛋白有20余种,其中包括了ApoAⅠ、AⅡ、AⅣ、B48、B100、CⅠ、CⅡ、CⅢ0-2、D、E、F、G、H(又称β2糖蛋白)、富含脯氨酸蛋白和甘氨酸-丝氨酸蛋白等(下表)。
主要的载脂蛋白
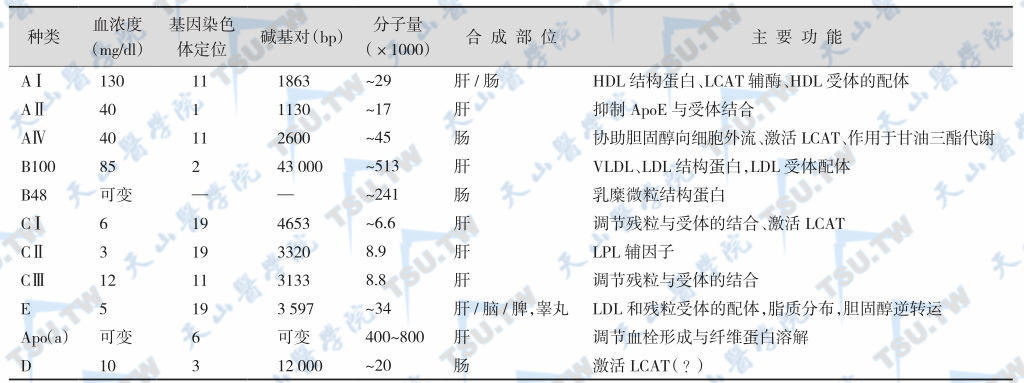
注:LCAT:卵磷脂胆固醇酰基转移酶;HDL:高密度脂蛋白;LDL:低密度脂蛋白;VLDL:极低密度脂蛋白;LPL:脂蛋白脂酶。
大部分载脂蛋白由肝脏合成,有11种载脂蛋白[如AⅠ、AⅡ、B48、B100、CⅠ、CⅡ、CⅢ、E、apo(a)等]的一级结构已阐明。载脂蛋白的功能有:
- 维持脂蛋白的结构;
- 作为酶的辅因子,如ApoCⅡ和ApoAⅠ是脂蛋白脂酶和卵磷脂胆固醇酰基转移酶(LCAT)的辅因子;
- 作为脂质的转运蛋白,如HDL中的ApoD使TG 和CE在HDL、VLDL和LDL之间的转运;
- 作为脂蛋白受体的配体而与受体特异性识别和结合,介导脂蛋白受体代谢途径,如ApoB100和ApoE是LDL受体的配体,ApoAⅠ是HDL受体的配体,等等。
脂蛋白的转化主要取决于其表层中的特异性载脂蛋白。研究显示,不少血脂谱异常症由载脂蛋白与受体结合功能异常所致,了解载脂蛋白在脂质代谢中的作用有助于理解脂蛋白代谢过程和脂质异常相关性疾病的发病机制。
脂蛋白受体调节脂蛋白代谢
脂蛋白受体包括LDL受体、LDL受体相关蛋白(LDL receptor-related protein,LRP)、gp330受体和VLDL受体,其基本结构相似而功能各异。
低密度脂蛋白受体
低密度脂蛋白受体(LDL受体)是一种分子量为160kD的糖蛋白。多种细胞表面都有LDL受体表达,尤以肝细胞明显。LDL受体在LDL、乳糜微粒残粒、VLDL、VLDL残粒、IDL和HDL1的摄取过程中发挥重要作用。细胞通过LDL受体摄取上述脂蛋白而获得胆固醇。LDL受体变异可导致脂代谢紊乱(遗传性家族性高胆固醇血症)。
低密度脂蛋白受体相关蛋白
低密度脂蛋白受体相关蛋白(LRP)是一种膜受体,由1个515kD的氨基端细胞外区和1个85kD的跨膜区构成。LRP与富含ApoE的乳糜微粒残粒和VLDL残粒呈高亲和力结合,通过与LPL和肝脂酶之间的相互作用调节肝细胞对脂蛋白残粒的结合与摄取。低密度脂蛋白受体相关蛋白-1(LRP1)主要转运细胞内的胆固醇,通过内吞作用(endocytosis)转运(胞转作用transcytosis)40多种不同结构的配体,因此LRP1是脂质通过血脑屏障、细胞质与核膜的最重要转运体。研究发现,LRP1还是清除淀粉样β肽(amyloid β-peptide,Aβ,导致Alzheimer病)的重要转运体;LRP1功能障碍是引起Alzheimer病的关键因素。
gp330受体
gp330受体是1种分子量600kD的蛋白质,又称为Heymann肾炎抗原。该受体在肾脏的近曲小管和脑室管膜细胞均有表达。尽管gp330受体可与含ApoE的脂蛋白和LDL结合,但其在脂代谢中的具体作用尚不清楚。
VLDL受体
VLDL受体与LDL受体非常相似,但具有1个第8配体结合重复序列,分子量130kD,主要存在于肌肉、脂肪和大脑组织中。该受体可与含ApoE的脂蛋白结合,其在脂代谢中的作用有待进一步探讨。
糖蛋白330受体
糖蛋白330(gp330/megalin)受体(600kD)为Heymann肾炎抗原,是组装LDL受体复合物的原料,一般认为与含ApoE的脂蛋白代谢有关。
ApoE受体2
为LDL受体家族的新成员,主要在脑组织中表达,是中枢神经脂代谢的关键受体。
清道夫受体
又称乙酰化LDL受体(acetyl-LDL receptor),目前已经发现5种类型(A~E),而所有的清道夫(scavenger)受体都具有与氧化型LDL和修饰型LDL结合的功能。可与结构修饰了的LDL(如乙酰化LDL或乙酰乙酸化LDL)相互作用,但不与天然LDL结合。清道夫受体还可清除体内的微生物、衰老细胞、变性的LDL和其他脂蛋白。血小板表达B类清道夫受体——CD36和SR-BI,它们主要参与血小板的血栓栓塞病理反应,与高胆固醇血症患者的急性心血管事件密切相关。血小板CD36是内源性氧化型磷脂酰胆碱(endogenous oxidized choline phospholipids)家族中的成员,可激活血小板功能,促进血栓形成,并同时降低HDL-C。血脂异常时,血清CD36增高。血小板SR-BI也有类似作用,但氧化型高密度脂蛋白(oxidized high-density lipoprotein)是SR-BI的配体,可抑制血小板的上述功能。
脂酶调节脂蛋白代谢
脂蛋白脂酶
脂蛋白脂酶(LPL)由448个氨基酸残基组成,分子量50kD。脂肪细胞、骨骼肌细胞、心肌细胞和巨噬细胞均可合成LPL。LPL从上述细胞分泌后,即被转运到毛细血管的内皮细胞表面,在此参与血浆中乳糜微粒和VLDL的分解代谢,调节TG的水解,释放非酯化脂肪酸供组织利用。LPL具有肝素、脂质、ApoCⅡ和LRP等4种物质的结合位点及1个催化位点。LPL是一种酯化酶,具有TG水解酶的活性及少部分磷脂酶活性。ApoCⅡ刺激LPL活性。LPL变异和ApoCⅡ缺陷导致高TG血症。
肝脂酶
肝脂酶是一种磷脂酶,具有TG水解酶的活性。分子量为53kD,由477个氨基酸残基组成。肝脂酶由肝细胞合成并存在于肝脏内皮细胞中。从肝脏分泌后,肝脂酶被转运至肾上腺、卵巢和睾丸的毛细血管内皮细胞。雄激素可增强肝脂酶的活性,雌激素则对此酶的活性有抑制作用。肝脂酶在脂蛋白代谢中的作用是多方面的。首先,它参与乳糜微粒残粒最终处理过程中的TG水解,还可能参与乳糜微粒表面过多磷脂的水解。其次,将IDL转化为LDL。此外,肝脂酶还可去除HDL2中的TG和磷脂,使HDL2转化成HDL3。ApoE是肝脂酶的辅因子,可促进其对TG和磷脂的水解。肝脂酶缺陷引起脂蛋白残粒、IDL和HDL水平的改变。
卵磷脂胆固醇酰基转移酶
卵磷脂胆固醇酰基转移酶(LCAT)由416个氨基酸残基组成,分子量为46.1kD,共有4个糖化位点。主要作用于小颗粒HDL和少数LDL,将其中的卵磷脂2位上的长链脂肪酸转移至胆固醇,生成溶血磷脂酰胆碱和胆固醇酯。体内大多数脂蛋白中的胆固醇酯都是在LCAT的作用下形成的。LCAT缺陷导致血浆游离胆固醇升高和胆固醇酯降低。
食物摄取影响脂代谢
饮食中的脂肪可提供足够的脂肪酸,但当饮食中糖类与脂肪的比例升高时,则刺激肝脏和脂肪组织中的脂肪酸合成。由于乙酰CoA是在线粒体内产生的,因此必须先与乙酰草酸缩合成枸橼酸盐才能进入细胞质,然后再转变成乙酰CoA和乙酰草酸。经过一系列反应,由8个单位的乙酰CoA共同缩合成棕榈酸。较长的脂肪酸(如硬脂酸和油酸)是通过增加棕榈酸链的长度而合成的。体内不能合成某些人体必需的多不饱和脂肪酸(如亚麻油酸和亚麻酸),必须从食物中补充。这些脂肪酸是机体许多特殊功能(如合成前列腺素)所必需的。
胃和近端小肠吸收食物中的脂肪。TG被水解为非酯化脂肪酸和少量的甘油单酯及甘油二酯,胆固醇酯水解成游离胆固醇,磷脂则转变成溶血磷脂酰胆碱。胆酸盐分子团扩散后,可溶解部分非水溶性脂质,有利于脂质转运至肠上皮细胞,并由此摄入细胞内。胆酸还可激活参与胆固醇水解的胰脂酶。长链脂肪酸主要在十二指肠和空肠上段的上皮细胞被摄取,再次酯化为TG及脂蛋白,随后经肠系膜淋巴管和胸淋巴导管进入体循环。10碳以内的中链脂肪酸无需经过酯化即可被吸收进入门脉系统,并在肝内廓清。胆酸在回肠经重吸收后进入门脉系统,被肝脏摄取。体内脂蛋白代谢过程见下图。
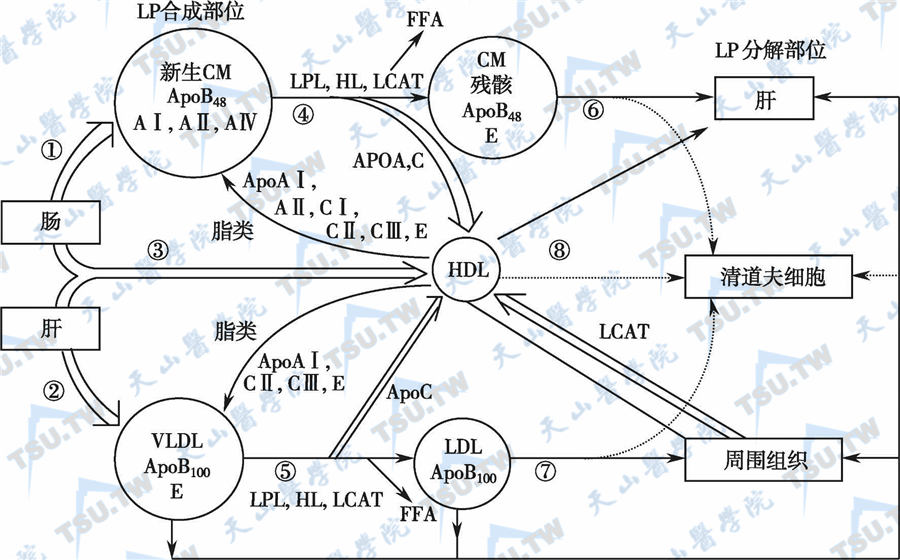
脂蛋白代谢示意图
注:LP:脂蛋白;CM:乳糜微粒;LPL:脂蛋白脂酶;VLDL:极低密度脂蛋白;HDL:高密度脂蛋白;LDL:低密度脂蛋白;LCAT:卵磷酯胆固醇酰基转移酶;HL:肝脂酶;FFA:游离脂肪酸。虚线示合成代谢,实线示分解代谢。
