
从显微镜下分辨出血液中的淋巴细胞已有100多年的历史,可是过去一直以为它们都是终期细胞,后来发现,外周血的淋巴细胞可能发生逆转,重新变成形态学彼此不同的幼稚细胞。曾经有一个阶段误认为淋巴细胞能够转化成粒细胞、单核细胞、巨噬细胞、成纤维细胞及其他细胞,后来才知道其各有起源。1935年,McMaster和Hudack第一次证实了抗体产生于淋巴结。与这些研究同时进行的是Fagraeus的经典研究,其工作显示了浆细胞产生抗体的直接证据。后来Coons等人采用免疫荧光技术得出结论,证实浆细胞内的确产生抗体。仍然存在的疑问是浆细胞是否起源于淋巴细胞。10年后由Harris等人通过细胞超微结构,证实了淋巴细胞转化为产生抗体的浆细胞的过程。同时,Nowell明确提出小淋巴细胞是具有增殖能力的细胞,能够转化原始淋巴细胞。
淋巴细胞为具一群有异质性的细胞,在细胞起源、生命周期、淋巴器官内的定居区域、表面结构和功能等诸多方面差异很大。虽然可以从一些形态结构,如细胞大小、颗粒和核质比例,来区别淋巴细胞群,但是这些指标不能证明它们的起源和功能。目前,临床实验室中应用的最精确和最有效的方法是利用单克隆抗体和流式细胞仪,根据淋巴细胞膜上的表面标记来确定淋巴细胞亚群。特异性抗原或者有丝分裂原等物质都可以刺激淋巴细胞增殖。非特异性的有丝分裂原引起的淋巴细胞活化是向原始细胞转化。通常有丝分裂原指的是任何可以引起细胞有丝分裂的物质。
淋巴细胞
淋巴细胞形态
一、光学显微镜
外周血中的多数淋巴细胞是小淋巴细胞(小于或等于10μm),大淋巴细胞少一些。有些大淋巴细胞被称为大颗粒淋巴细胞(large granular lymphocytes,LGLS),胞质中含有嗜天青颗粒。确定淋巴细胞直径时,会因测量细胞方法不同出现一些变化。在瑞-吉染色的血涂片中,淋巴细胞的核是深紫蓝色,是由染色质聚集而形成的,通常为圆形或有轻度凹陷。普通染色方法不能观察到淋巴细胞的核仁,但在组织切片中有时可看见一到两个核仁。
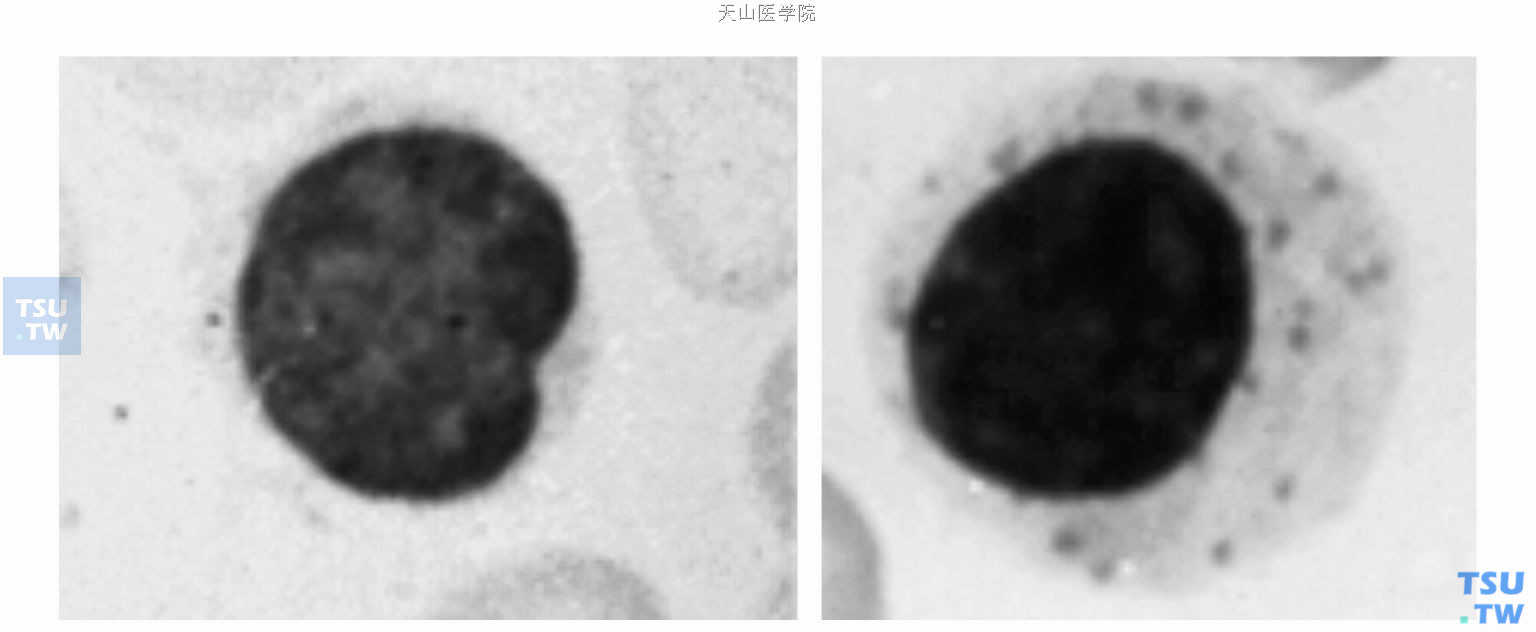
小淋巴细胞和大颗粒淋巴细胞
二、透射电子显微镜
小淋巴细胞(6~9μm)呈现出一种光滑、双层的细胞膜结构,细胞膜上有少量的微绒毛,在细胞表面偶尔可见一些泡状结构,内有膜包裹着的胞质。小淋巴细胞胞质中往往没有细胞器,高尔基体也很小,位于细胞核凹陷处。在切片的合适断面上可以见到高尔基体中有一两个中心粒,相互垂直。淋巴细胞不易观察到内质网,但是可见许多游离核糖体,偶尔可见核糖体群。典型的线粒体很常见,但溶酶体少见,偶尔可见不明意义的致密体(dense bodies)。
淋巴细胞的细胞核由一双层膜相结构包绕,双层膜在核孔处融合,膜周围聚集着大量的异染色质,淋巴细胞核内的异染色质不很多。这些异染色质的缝隙间含有常染色质、核糖体样颗粒和原纤维。通常可见一个核仁,中淋巴细胞因为胞质量多,体积稍大一些,核染色质疏松,有些小的块状异染色质黏附于核膜上,高尔基体也比小淋巴细胞的发达一些,胞质中也含有大量的多聚核糖体和一些内质网,大多平行于核膜分布。
原始淋巴细胞体积常常更大(8~12μm),其核染色质松散,有一个巨大的核仁,清晰可见,核占据了整个细胞的1/3。胞质含有大量多聚核糖体,但内质网少见。淋巴细胞和其他体细胞一样,有丝分裂期可以分辨出44条常染色体和2条性染色体。
三、扫描电子显微镜
以前在扫描电镜观察人类外周血中的淋巴细胞时,根据淋巴细胞表面形态将其分为两大类:一种表面光滑;另一种表面呈毛绒状,覆有大量微绒毛。曾认为前者是胸腺来源的(T)细胞;后者是骨髓来源的(B)细胞。后来的研究结果表明,淋巴细胞表面的一些特征是制备方法和细胞所处的不同功能阶段造成的。在不同有丝分裂原刺激下的淋巴细胞具有不同的微绒毛,此时的微绒毛与淋巴细胞的来源无关。当淋巴细胞处于它们最合适的微环境时,所有淋巴细胞的表面都是光滑的。休眠淋巴细胞往往表面光滑,微绒毛的出现往往是由环境刺激和细胞表面受体造成的。毛细胞白血病和脾边缘带淋巴瘤细胞的微绒毛均较长,是电镜下诊断相应疾病的依据之一。
淋巴细胞的体外转化
特异性抗原或者有丝分裂原等物质都可以刺激淋巴细胞引起反应。非特异性的有丝分裂原引起的淋巴细胞活化是原始细胞转化。通常有丝分裂原指的是任何可以引起细胞有丝分裂的一些物质。最常用的是植物血凝素(PHA)。其中一些血凝素可以特异性地与T或B细胞结合,另一些则与两种细胞都起作用。植物血凝素作用于T淋巴细胞。商陆素(PWM)可以在T细胞缺乏时引发B细胞分泌免疫球蛋白。有丝分裂原刺激的淋巴细胞转化要求有丝分裂原结合到适当受体上,这是淋巴细胞活化十分关键的一步。
淋巴细胞活化的第一步是适当的受体相互交联。在受体结合后几秒到几分钟的时间内,相继发生了如下反应:一价阳离子(Na+和K+)流入细胞;膜相关甲基转移酶激活,催化甲基的转移。同时,细胞磷脂的成分之一磷脂酰肌醇合成量增加。PHA与人淋巴细胞结合后的1~5分钟内钙离子流入在活化中起重要作用,可能与关键酶或蛋白的磷酸化有关。细胞在结合了有丝分裂原后的几个小时内,蛋白合成增加,在48~72小时达到高峰。多种蛋白在短时间内大量合成。在这期间淋巴细胞的RNA合成也增加了。RNA在48小时内翻了一番,细胞表现出明显的形态学变化。RNA合成增加标志着淋巴细胞的活化,合成增加发生于36小时,48~72小时达高峰。
被PHA活化后的淋巴细胞变成大体积的淋巴细胞(10~20μm),由于RNA含量增加胞质呈深蓝色。通常有丝分裂象出现在40小时,72小时达高峰。通过放射自显影分析可见嗜碱性颗粒集中在细胞核周围。细胞核变大,呈纤细网状。早期可见细胞核仁的变化,包括颗粒变大、聚集,最后出现在核内染色质中。胞质的变化包括胞质量急剧增加,核糖体聚集增加和高尔基体变大。PHA刺激人类外周血淋巴细胞转化后获得一个非常精细的囊泡系统,包括细胞内泡,多泡结构和致密小体。囊泡系统不同成分都起源于胞内自身的小囊泡,参与摄取、转运、储存和消化胞内物质的作用。形成密集的溶酶体泡需要明显的PHA刺激因素。PWM诱发细胞转化的峰时在72小时,50%~60%的淋巴细胞可变为转化细胞。在组织培养中可以看到三种细胞:未转化的小淋巴细胞、形态学上类似原始浆细胞的原始细胞和不成熟的浆细胞。浆细胞细胞核偏心,染色质聚集在一起,高尔基体发育旺盛,胞质出现大量内质网,这些改变是PWM刺激引起淋巴细胞大量合成的免疫球蛋白的形态学证据。
淋巴细胞运动
淋巴细胞运动时往往细胞先伸出了一只伪足,伪足逐渐从胞体上分离。细胞核被推向狭窄的胞质中的另一边,像一面手镜。细胞的前端是细胞核,大部分胞质被拖在后面,形成镜子的手柄。胞质形成的尾部被称作“尾足”[由希腊语ura(尾巴)和pod(足)而来]。它们有0. 125~0. 2μm宽和0. 8μm长,其上漫布绒毛和微丝。淋巴细胞运动的速度平均为20μm/min。一般来说,原始淋巴细胞的运动速度要快于小淋巴细胞,刺激诱导淋巴细胞向原始细胞转化可使细胞的运动增强。淋巴细胞的尾足不仅仅与运动有关,还是与其他细胞,如巨噬细胞等,相互作用的部位。在淋巴细胞混合培养中,淋巴细胞可以接近巨噬细胞,通过尾足与它们建立联系。T淋巴细胞也可通过尾足聚集成玫瑰花簇黏附至巨噬细胞上。淋巴细胞和巨噬细胞只有当暴露给抗原后,才能形成稳定的聚集簇。因此,尾足有可能在抗原结合和识别过程中起一定作用。T细胞簇可以刺激细胞分化,淋巴细胞发现尾足是细胞激活的形态表现。淋巴细胞的运动与其他细胞的运动有很大的不同。淋巴细胞的运动很平稳,保持为手镜形状,而原始粒细胞的运动像龙卷风样扭转,单核细胞系的细胞在运动中不停地改变形状和方向。
尽管很多人认为尾足是T淋巴细胞的特征,实际发现B淋巴细胞也能在抗免疫球蛋白抗体的刺激下形成尾足。尾足形成往往是表面抗原被覆盖之后,呈帽状完全覆盖在尾足上,最后内吞入细胞。尾足也被认为是外界物质内吞入胞的位点。尾足可以有胞质内的一切细胞器,包括高尔基体、线粒体、微丝和微管。
浆细胞
Harris及其同事研究动物免疫细胞的超微结构,他们分离了合成抗体的单个细胞。发现当细胞仍然保持为“淋巴细胞”的形态时,就可以检测到抗体的产生,这些细胞不含有内质网。这些细胞可以从非活化的淋巴细胞进一步分化,产生大量的游离多聚核糖体。细胞分泌抗体的能力大小依赖其内质网的发育程度。在分化早期,内质网少而分散,随后,内质网层的长度增加并且平行排列,逐渐充满全部胞质,形成独特的洋葱皮样结构。高尔基体同时增大,有淋巴细胞形态的细胞常产生IgM;而有浆细胞形态的细胞常产生IgG。但是,产生IgM抗体的细胞和产生IgG抗体的细胞不存在一条严格的分界线。1875年,Waldeyer首次应用“浆细胞”一词,然而,他的描述实际包括了几种形态的细胞。1881年,Unna在观察一例狼疮患者的细胞时重新定义浆细胞,强调其胞质嗜碱性的特点。接下来的报道中包括了甲基绿和派洛宁染色细胞的图片,一些细胞很容易识别出浆细胞的特点。1895年Marschalko在讨论Unna所描述的浆细胞形态时,重点提出了细胞核内的染色质成一定角度聚集的特点(车轮状)。细胞核位于偏心位置,可用作识别浆细胞的严格指标。Pappenheim为浆细胞核的形态学特点起名为radkern。Downey和Michels先后综述了有关浆细胞的特征,这期间,浆细胞是否属于正常的组织成分一直是许多争论的焦点。它的起源也存在争议,但是一些著名的研究者认为浆细胞起源于淋巴细胞。
1937年Bing和Plum的临床观察使浆细胞的研究进入一个新的阶段,他们发现高免疫球蛋白血症与浆细胞密切相关。之后,Bjornboe和Gormsen证实免疫兔的体内抗体的产生与脾脏中浆细胞的大量增殖有关。Fagraeus提出动物之间产生抗体能力可能与它们的浆细胞数量,特别是不成熟浆细胞的数量有关,认为成熟的浆细胞不得不“经历它们巨大的功能性强度阶段”。
Coons为浆细胞即是产生抗体的细胞提供了无可争辩的依据,他将免疫荧光技术应用于免疫学研究。在脾脏的红髓、淋巴结的髓索及免疫动物的灶状细胞聚集部位检测到包含抗体的浆细胞。
浆细胞在形态学上很容易与其他类型的细胞区分开。细胞呈球形或椭圆体状,5~30μm大小。胞质丰富,嗜碱性,多呈深蓝色,可以有颗粒。浆细胞核周有一核周淡染区,核旁有高尔基体。浆细胞的胞核相对体积较小,胞核呈圆或卵圆,有浓聚的染色质,呈车轮状排列。
有些浆细胞胞质染色成为红色或紫色而非蓝色,被Undritz称之为“火焰”状的浆细胞,这是由于大量内质网聚集,其中含有高碳水化合物的Ig。电子显微镜下的浆细胞核是双层膜状结构,外层膜覆盖核糖核蛋白的颗粒,并且延伸为胞质内质网。高尔基体由囊泡和小管组成,紧挨着核,中间包绕中心粒。一些微管以中心粒为中心,呈放射状分布,许多线粒体分布于内质网层之间。具有丰富且排列有序的内质网是浆细胞超微结构的一个重要特点。内质网的一侧布满核糖体颗粒,平行排列成层状的膜结构。成熟浆细胞的内质网充填整个胞质。内质网有时由于颗粒成堆而膨胀,成为胞质包涵物,称为拉塞尔(Russell)小体。拉塞尔小体由免疫球蛋白组成,但常常很难证实。有时可在细胞核内看到拉塞尔小体(核内包涵物)。在某些情况下,浆细胞内大量的同源性物质充填整个细胞,使其染为灰色,称为储积细胞。在电子显微镜下均为内质网膨胀所致。火焰状细胞是储存了免疫球蛋白的早期细胞,是免疫球蛋白分泌被阻滞后形成的。Mott细胞也被认为是具有多个拉塞尔小体的浆细胞,也是因为免疫球蛋白分泌完全或部分被阻滞,内质网局部膨胀所致。
原始浆细胞是最不成熟的浆细胞,核浅染,染色质疏松,可见一个较大的核仁。内质网稀少,胞质内充满一簇簇的多聚核糖体。当细胞成熟时,染色质凝聚成块状,内质网越来越丰富并且排列有序,大量内质网聚集在形态学上称为“动浆”。淋巴细胞成熟为浆细胞时必然经历这个中期阶段。
浆细胞经常可见不典型的形态,一些浆细胞可以表现为淋巴细胞样的形态,同时有一个不寻常的粗面内质网。这种中间形态细胞可以称为淋巴细胞样浆细胞或浆细胞样淋巴细胞。在浆细胞瘤和患有高免疫球蛋白的免疫疾病的患者血中常见这类细胞。在病毒感染的患者血中也可见到类似的细胞(Turk细胞)。此外在传染性单核细胞增多症和部分健康人血中也能见到类似细胞,未成熟浆细胞很像PHA刺激转化的细胞,核很大,染色质呈细网状,胞质内的内质网相对量少,可见许多核糖体和多聚体,因此很难划分这些细胞。另有一些浆细胞胞质中含有空泡或是面条样的内涵物。
上皮细胞
超微结构下,上皮细胞有不同的细胞亚群,反映了它们的来源不同。上皮细胞曾分为许多不同的种类,如网状组织、合胞体、上皮组织和鳞状细胞。皮质中的上皮细胞胞质有25μm长,与树突状上皮细胞相同,这一特征将它区别于胸腺实质区的其他细胞。它们都具有桥粒和张力细丝,张力细丝是纤细结构,长度0. 1~0. 3μm不等。通常成群分布于核周,在相邻上皮细胞的树突状连接中可见桥粒。有卵圆形的核,均匀分布的染色质和明显的核仁。上皮细胞通过桥粒相连,形成网状结构,包围着血管、小梁和囊内壁。在上皮间及小梁和血管周围有一种与毛细血管基底膜完全不同的基底膜,上皮网状结构就这样充满了淋巴的空隙。
在皮质中最多见树突状上皮细胞,没有分泌颗粒,可能不参与胸腺激素的分泌,通过细胞相互作用来指导淋巴细胞分化。这些相互作用由主要组织相容性复合体的基因表达产物进行调节。已经证实细胞的Ⅰ类和Ⅱ类抗原分子在小鼠和人类表达。皮质上皮细胞上,Ⅱ类抗原要比Ⅰ类抗原密集,但在髓质上密度相同。免疫组化技术和电镜分析得知这些分子位于具有桥粒和张力细丝的上皮细胞上。Ⅱ类抗原可在14天小鼠胚胎中的胚胎上皮细胞中检测到。在完全缺乏成熟T淋巴细胞的动物裸鼠身上,胸腺基质中完全缺乏Ⅱ类抗原。
外层皮质内的胸腺细胞和上皮细胞间相互作用,可形成所谓的保姆细胞。由大集落淋巴细胞周围包绕上皮细胞。髓质中的上皮细胞没有很长的树突形状而被称为上皮状或平板上皮状细胞。它们常为多型的,具有黑色颗粒,可能具有分泌功能,另外一种上皮细胞即鳞状上皮也广泛的存在于髓质内。这些细胞可形成Hassall囊并且具有大量的张力细丝和角质透明蛋白。
Hassall囊常为实体或囊状。20~50个上皮细胞紧密排列,形成的固相结构的小体,中央区的细胞发生了变形而形成囊状。这些囊的表面有内皮细胞覆盖,在一些固化的囊中有时也有碎屑和变形的细胞,角质十分显著,与皮肤中的也很相似。深皮质内的上皮细胞与髓质中的表型不同,最终可以角化。
巨噬细胞
胸腺组织中的巨噬细胞常在血管周围出现。一些胸腺细胞围绕巨噬细胞形成玫瑰花状。胸腺巨噬细胞诱导胸腺成熟。一般认为巨噬细胞来源于单核细胞,胞体较大,直径15~40μm,胞核成网状,往往偏位,胞质丰富,为淡灰蓝色或者浅红色。淋巴结的巨噬细胞内含有大量淋巴细胞或者其他吞噬物的碎片,起清除凋亡细胞的作用。
树突细胞
除吞噬细胞外,树突细胞(DC)也具有抗原递呈功能。DC细胞体可以伸出弯曲的胞质枝杈,或称树突。DC存在于淋巴和非淋巴组织,如皮肤、肺、内脏黏膜。它们能够有效的递呈抗原并提供T、B细胞的辅助信号,包括高浓度的MHCⅠ、Ⅱ类分子和许多黏附分子。DC在其整个生命过程中常改变表型,通过外形或个体发生学鉴定它的存在很困难。它们是如何从单核-吞噬细胞分化出来的尚不清楚。具有吞噬活性被认为是粒单细胞的标志。DC缺乏趋化活性,故被认为是另一类细胞。但近期研究表明,在骨髓和外周血存在着DC的CD34+前体细胞。体外培养发现巨噬细胞和DC克隆有各自的单独干细胞来源,表明这两种细胞是不同的。在外周血单个核细胞发现一种更为成熟的有两种潜能的前体细胞。外周血DC前体细胞接受不同的信号可以分化为巨噬细胞或DC。例如,IL-4就促进DC分化。在脐带血,CD34+前体细胞能分化为DC,此DC是CD1a+,表达CD13、CD4、DR和不同量的CD14。可能有不止一种的分化途径使有淋巴细胞和无淋巴细胞存在的不同区域产生DC。
第一条皮肤分化DC的途径是由GM- CSF、TNF-α刺激CD34+前体细胞产生,这类细胞表达皮肤的白细胞抗原(CLA)。
第二条途径产生髓系树突细胞(MDC),来自CD34+的骨髓前体细胞。采用不同刺激方法有两条潜在的分化途径,分别产生MDC或巨噬细胞。GMCSF和TNF-α刺激产生MDC,而M- CSF刺激产生巨噬细胞。MDC短暂表达CD14,但与巨噬细胞同样有其他标志如CD13、CD33和CD36。
第三条途径产生淋巴样DC,其前体细胞先是在胸腺中出现,为CD4弱阳性细胞,缺乏其他髓细胞标记,无需GM- CSF诱导分化。外周血存在其中间前体细胞CD4+、CD11-细胞,在IL- 3存在的情况下分化成成熟DC。这些细胞形态上类似原始浆细胞。
不管其确切的发育途径是什么,通过与GMCSF、TNF-α、IL-4、干细胞因子和CD40配体共同培养,外周血的某些前体细胞能发育成DC。从各种途径来的成熟细胞通过不同黏附分子和归巢受体,以及标记和转移途径,到达特定部位安居下来。虽然特殊标记和转移途径可作为不同DC的标志,但实际它们都共有一些特征,像DC的形态学、提呈抗原的能力、缺乏趋化性、可以激活T细胞增殖等特征。DC另一个非常重要的特征是从非淋巴组织将抗原输入淋巴管运至淋巴结。在淋巴结中,外界抗原的最终归途是T细胞依赖区域,DC提呈抗原给T细胞,激活并启动免疫反应。
滤泡状树突细胞(FDC)
在淋巴结的滤泡和胚系生发中心可以找到滤泡状树突细胞(FDC),FDC识别滞留于此的抗原。抗原滞留还需抗体Fc片段和补体参与。与被巨噬细胞捕捉的抗原相比,抗原于此滞留很长时间。从形态上看,FDC有不规则的核,细胞质很少,丝状树突使细胞像太阳光芒般的形状。在其表面表达Fc和C3受体和Ⅰa抗原。免疫后,抗原传递至局部淋巴结的淋巴窦内。这些传递细胞被称为抗原转运细胞(ATC),抗原转运细胞形成运输链。ATC携带抗原到FDC。FDC结合淋巴滤泡,这一相互作用形成了滤泡网。滤泡网又称抗原滞留网(ARR),ARR发育需B细胞的参与。ARR是FDC在滤泡的亮区形成的三维网,电镜发现有两种FDC:丝状突的FDC和珠状突的FDC。前者可以转变为后者。二次免疫后,FDC形成球形颗粒称为免疫复合物包被小体(iccosomes),至细胞生发中心解离。FDC可通过补体依赖片段C3d和C3dg与FDC复合物结合,传导共刺激信号到B细胞。结合到B细胞上补体受体(CR2,CD21)。CR2或CD21是CD19/CD21/TAP1复合物的成分,是B细胞抗原受体。FDC相关复合物与B细胞受体和CD19/CD21/TAP1共同受体结合,为B细胞激活提供了两个信号。CD21以长的和短的两种形式存在。FDC选择性表达CD21长异构体,而B细胞表达短的异构体,FDC的CD21最可能参与复合物形成,IL- 2加强FDC依赖的B细胞刺激。
FDC可阻滞逆转录病毒颗粒。感染的FDC携带病毒,增强了对CD4+T细胞的感染性。FDC的抗原递呈功能会逐渐丢失并破坏。通常DC对HIV感染很敏感,可能是体内HIV感染最早的细胞。它们分布在皮肤和黏膜表面。DC携带病毒到淋巴结内,递呈抗原给T细胞,在相互作用中进一步感染T细胞。
并指状树突细胞(IDC)
T细胞依赖区存在一定数量的IDC,胞质清晰,胞核不规则,胞质形成一些长的树突,可与DC作用。IDC趋化能力与吞噬细胞不同,缺乏酸性磷酸酶和非特异性颗粒,吞噬细胞活性弱。它们表达MHCⅡ类抗原,但不表达免疫球蛋白Fc段受体和补体,IDC CD83和S- 100b蛋白阳性。从其他部位(皮肤和脾脏)来的IDC和DC有共同特征。IDC很可能和DC属于同一系统,外周非淋巴器官的DC移至T细胞依赖区就成为IDC。
黑热病原虫进入皮肤后,可在淋巴结的T细胞区IDC中发现,可能是皮肤DC摄取原虫移至淋巴结,成了IDC。皮肤过敏的病人可以在局部淋巴结的一些IDC中发现Birbeck颗粒。T淋巴细胞的标记CD12强染色,支持淋巴细胞从皮肤移至局部淋巴结T细胞区的假说。外源性淋巴细胞将抗原转移给DC并非只是皮肤DC的特性,肠道DC也有相似的功能,可以将口服抗原移至局部淋巴结。
朗格汉斯细胞
这些细胞首先由朗格汉斯(Langerhans)在表皮发现,是一种多核巨细胞。特殊染色可发现其有对重金属离子的亲和力,并可摄取细胞的左旋多巴、多巴胺和去甲肾上腺素。其超微结构首先由Birbeck发现,他还发现了以他的名字命名的特异性颗粒。朗格汉斯细胞有数十个排列规矩的细胞核、胞质不规则,胞质中有微丝,多囊体和独特的Birbeck颗粒,此颗粒杆状,有Z形条纹。真皮淋巴管和边缘的淋巴结的副皮质可以见到这种细胞,表明这些细胞是可运动的。
脐带和成人外周血有CD34+朗格汉斯前体细胞,这类CD34+细胞仅仅会分化为带Birbeck颗粒的朗格汉斯细胞。成熟的朗格汉斯细胞表达颗粒性抗原。朗格汉斯细胞表达Ⅰa抗原、IgG、C3和Fc段受体,但是缺乏表面免疫球蛋白。它们在抗原提呈中比巨噬细胞效率更高,是皮肤中的主要抗原提呈细胞。敏感物质直接接触可激活朗格汉斯细胞。紫外照射可去除皮肤朗格汉斯细胞,阻断接触激活。朗格汉斯细胞作为辅助细胞的另一项功能是在混合淋巴细胞反应中协助简单的生化物质激活同种异型T淋巴细胞反应,引起T细胞增殖。
Vieled细胞
Vieled细胞的命名来自它们有很长的具有活性的遮盖物,遮盖物能达100nm厚,不含颗粒,可以延伸较远距离。Vieled细胞的核呈高度卷曲,胞质有许多颗粒。只有4%Vieled细胞有Birbeck颗粒,但是细胞表面之下都有大的空泡,用ATP酶检测为强阳性反应。在酶含量方面类似淋巴细胞,它们有Fc受体,但不能主动趋化。在接触激活后,朗格汉斯细胞出现于皮肤淋巴液中,淋巴管中Vieled细胞数量也增加。许多Vieled细胞移到淋巴结,定位于T细胞依赖区,在此位置细胞的作用像树突状细胞。一些Vieled可促进T细胞对有丝分裂原的反应。它们的功能包括转运抗原到皮质区,将其提呈给T淋巴细胞。因此,体内存在一个抗原提呈的细胞链,此链中有不止一种的细胞。只在皮质的深层发现具有Vieled细胞特征的细胞。
