
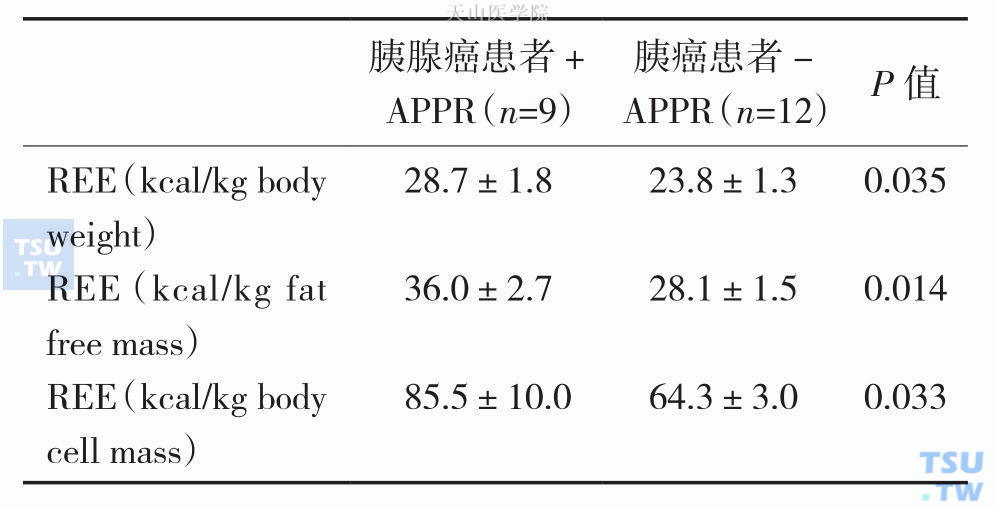
恶液质体重下降实际上是能量摄取严重下降,能量消耗不断增加,或两者同时存在所导致的结果,也即能量代谢的严重失衡所导致的一种临床恶性综合征。静息状态能量消耗(resting energy expenditure,REE)是衡量能量代谢的一个主要指标。一般指在进食后3~4小时处于休息状态时测定的能量消耗量。正常人静坐时REE占总能量消耗的70%,而肿瘤患者REE往往高于正常对照组,REE高低还与肿瘤类型相关,如肺癌和胰腺癌患者的REE显著升高,而胃癌和结肠癌患者的REE升高不明显。1994年Falconer J. S等对一组21例胰腺癌患者检测REE和炎症指标,不管是以体重、无脂肪组织质量,还是体细胞质量换算的REE都显著高于正常对照组,按体重计算的REE大约高33%(表3-1-1),同时发现伴有急性期蛋白反应(acute-phase protein response,APPR)(C反应蛋白>10mg/L)的胰腺癌患者REE也明显高于不伴APPR的患者(表3-1-2)。2001年HR Scott等对一组12例没有体重下降的非小细胞肺癌(non-small cell lung cancer,NSCLC)患者研究发现NSCLC患者REE约高于正常对照组15%(P<0.01),且与炎症反应程度密切相关(r=0.753,P<0.01),这反映了NSCLC患者体内的炎症反应对其REE增加产生重要的影响,且这些作用先于体重下降发生。
表3-1-1 胰腺癌患者与正常对照组REE(X±SD)
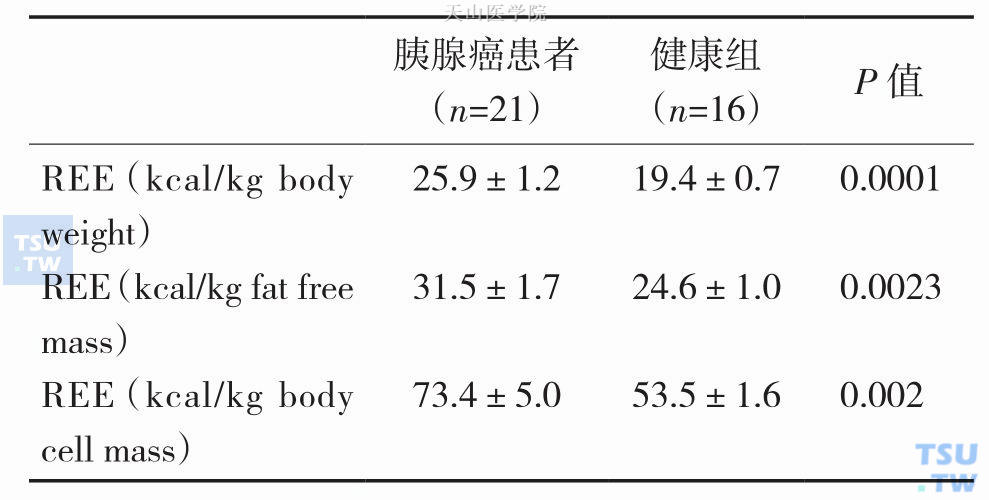
表3-1-2 胰腺癌患者伴有或不伴有APPR的REE (X±SD)
恶液质患者高能耗状态可能与以下三个方面有关:
炎症因子参与恶液质患者的能量消耗
细胞因子和炎症介质通常参与到肿瘤发生和发展的整个过程中,它们是肿瘤患者生存,生活质量以及治疗效果的关键因素。近年来研究进一步强调了肿瘤来源的细胞因子与恶液质的关系,相当多的证据揭示了IL-1、 IL-6和肿瘤坏死因子α(tumor necrosis factor-α,TNF-α),以及 myostatin/activin通路等发挥重要作用。REE显著升高胰腺癌患者常常伴随的急性期反应(acute-phase response,APR),在肺癌和胃肠道癌患者中APR程度与患者体重丢失率密切相关。1998年DC McMillan等对12例肺癌或胃肠道癌的男性患者进行为期12周的纵向研究发现体内总钾丢失量与C反应蛋白增加成正比(P<0.05),也与三头肌皮褶厚度下降密切相关(P<0.05),该结果提示癌症患者体细胞质量丢失率与慢性炎症反应程度密切相关。愈来愈多的证据表明在肿瘤恶液质情况下肿瘤来源的炎症因子信号干扰正常脂肪组织与其他器官之间信号交流,从而损害能量代谢平衡和脂肪组织和肌肉组织分解代谢平衡。
UCPs参与恶液质患者的能量消耗
某些恶液质患者高REE的主要原因是BAT和肌肉组织产热作用增强。BAT在动物和人的体温调节和能量平衡调节中发挥重要作用,但是一般来说成人只有很少的BAT。但是尸检结果显示80%恶液质患者肾上腺周围存在BAT,而正常对照组只有13%。BAT和肌肉组织产热源于UCP,与能量消耗相关UCPs主要包括UCP1、UCP2和UCP3。研究发现骨骼肌过表达UCP3转基因小鼠食欲旺盛而体重低于对照动物,并且脂肪组织明显减少,小鼠饥饿期里棕色脂肪组织的UCP3水平下降而骨骼肌的UCP3是升高的,这可能促进脂肪酸氧化。当荷瘤小鼠(MAC16结肠腺癌)体重丢失达24%时,棕色脂肪组织UCP1 mRNA水平明显升高,而骨骼肌UCP2和UCP3mRNA水平也明显升高;而在易产生恶液质的荷瘤小鼠(吉田腹腔积液肝细胞癌)也出现骨骼肌的UCP2和UCP3 mRNA水平明显升高。在这两种荷瘤小鼠模型中还发现骨骼肌UCP3 mRNA水平直接与血清游离脂肪酸水平相关。一种噻唑烷二酮类药物类降糖药曲格列酮可以选择性激活过氧化物酶体增殖因子激活受体-γ(peroxisome proliferator activated receptor-γ,PPAR-γ),而显著降低小鼠骨骼肌UCP2和UCP3 mRNA 水平,这提示PPAR-γ配体可降低恶液质能量消耗。因此,曲格列酮类药可能对于癌症患者有一定应用价值。这也表明增加UCP3表达可以提高能量消耗和组织分解代谢,其中的机制可能比较复杂。在大鼠恶液质模型中,UCP增加同时伴有2倍以上的血液游离脂肪酸升高,用烟酸降低高脂血症时也能降低比目鱼肌UCP3表达,而腓肠肌UCP3没有变化。而小鼠恶液质模型中UCP2 和 UCP3 表达增加并不与血液游离脂肪酸升高相关。有证据表明某些细胞因子和肿瘤脂肪动员因子(lipid mobilizing factors,LMF)可以增加BAT和骨骼肌中UCP 表达。
恶液质患者无效循环增加耗能
20世纪初,Warburg精巧实验揭示了人和动物肿瘤细胞在有氧的条件下仍进行活跃的糖酵解,这种现象被称为“Warburg效应”,并认为“Warburg效应”与肿瘤之间存在因果关系。糖酵解的增强与肿瘤的生长速度成正比,而与分化程度成反比。Warburg 效应是由于肿瘤细胞癌基因和抑癌基因变异导致代谢重编程的结果,重编程导致肿瘤细胞更能适应不利环境生存和促进增殖性合成作用。目前,Warburg效应作为肿瘤的能量代谢标志,已在多种类型的细胞中得到证实。肿瘤糖酵解除了低氧条件下获能外,更重要是为了获得增殖性合成生物大分子所需的大量构件分子。因此需要消耗大量的葡萄糖。研究发现肿瘤细胞糖酵解能力是正常细胞的20~30 倍,最高可达到正常的200倍。由于糖酵解产物是乳酸,因此肿瘤组织不断摄取葡萄糖而大量排出乳酸,这样在肿瘤患者肝脏与肿瘤之间形成类似Cori循环,即葡萄糖-乳酸循环(图3-1-7)。肿瘤细胞不断释放乳酸进入血液循环,在肝脏进行糖异生,产生葡萄糖又大量被肿瘤细胞摄取。从糖酵解能量产生和肝脏糖异生消耗能量来看是得不偿失的,在肝脏2mol乳酸合成1mol葡萄糖要消耗6molATP,这样就增加了葡萄糖和ATP的无效消耗,葡萄糖利用效率明显下降。研究证实转移性结直肠癌患者比对照组具有较高效率的葡萄糖-乳酸循环,这样通过Cori循环消耗能量达到 300kcal/ d。另外发现肿瘤乳酸水平可能与肿瘤转移和复发呈正相关,而与患者生存呈负相关。
由于肿瘤患者存在胰岛素分泌不足或胰岛素抵抗等导致肝脏糖异生通路异常活跃而消耗大量的能量。其他一些代谢物(甘油和生糖氨基酸等)也可进入肝脏进行糖异生而消耗能量,如肿瘤患者,尤其是恶液质状态下,脂肪组织和骨骼肌分解加强,产生大量的甘油和氨基酸进入肝脏糖异生。其中肝脏与肌肉之间的丙氨酸-葡萄糖循环也是一条重要的循环耗能通路(图3-1-8),2mol丙氨酸异生1mol葡萄糖时消耗6molATP,同时丙氨酸脱氨产生2摩尔NH3将会进入鸟氨酸循环通路合成1mol尿素消耗4molATP,这表明2mol丙氨酸通过丙氨酸-葡萄糖循环和鸟氨酸循环共消耗10molATP。此外,脂肪组织内脂肪分解增强除了释放甘油外还有游离脂肪酸(free fatty acid,FFA),游离脂肪酸除了氧化分解外还可以再酯化生成三脂酰甘油(triacylglycerol,TAG),这也就是TAG-FFA底物循环。这方面的循环耗能在恶液质患者和动物模型中的测定不多,其中在荷瘤小鼠模型中检测的TAG-FFA循环率显著高于非荷瘤小鼠组。
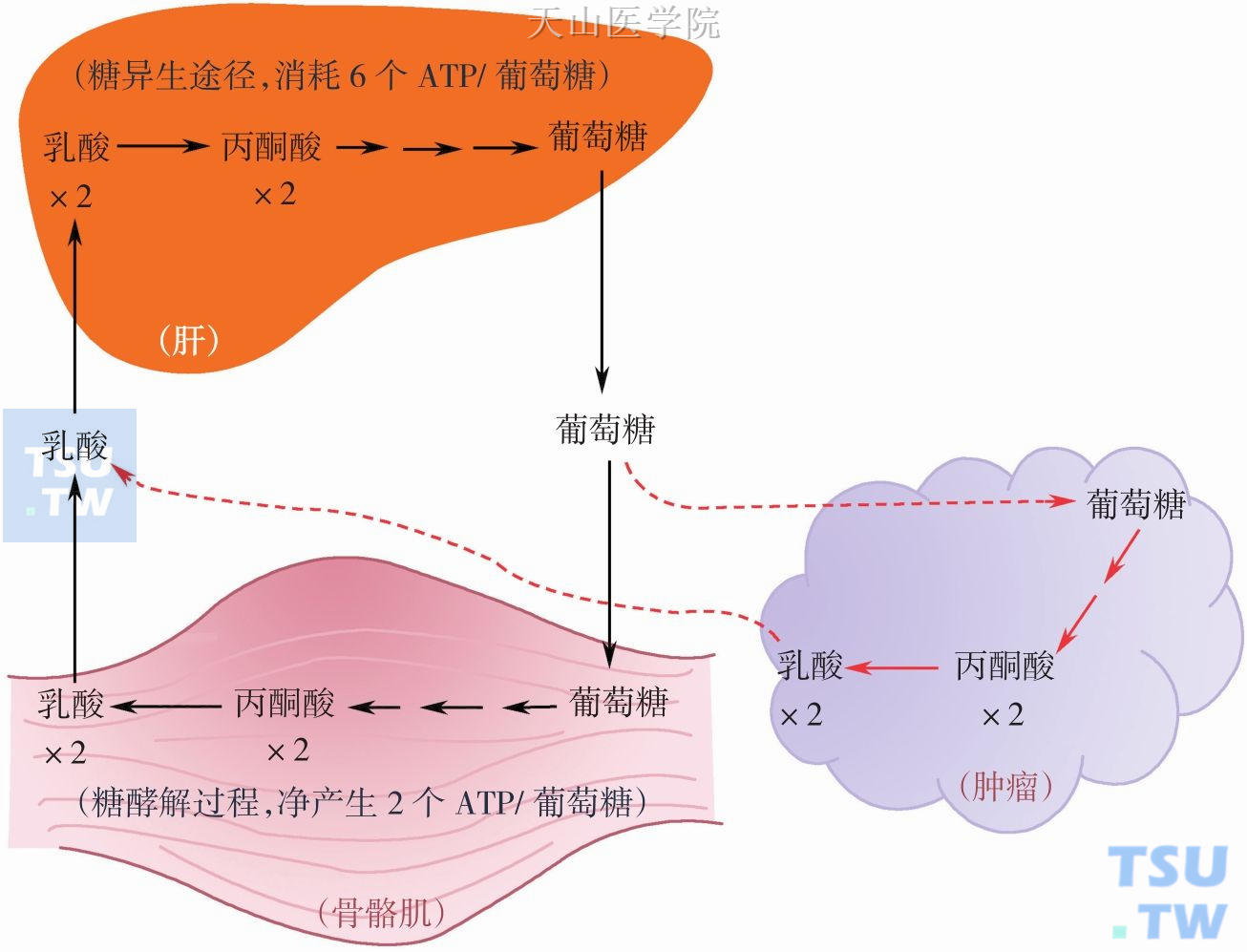
图3-1-7 肿瘤与肝脏之间的类Coris循环
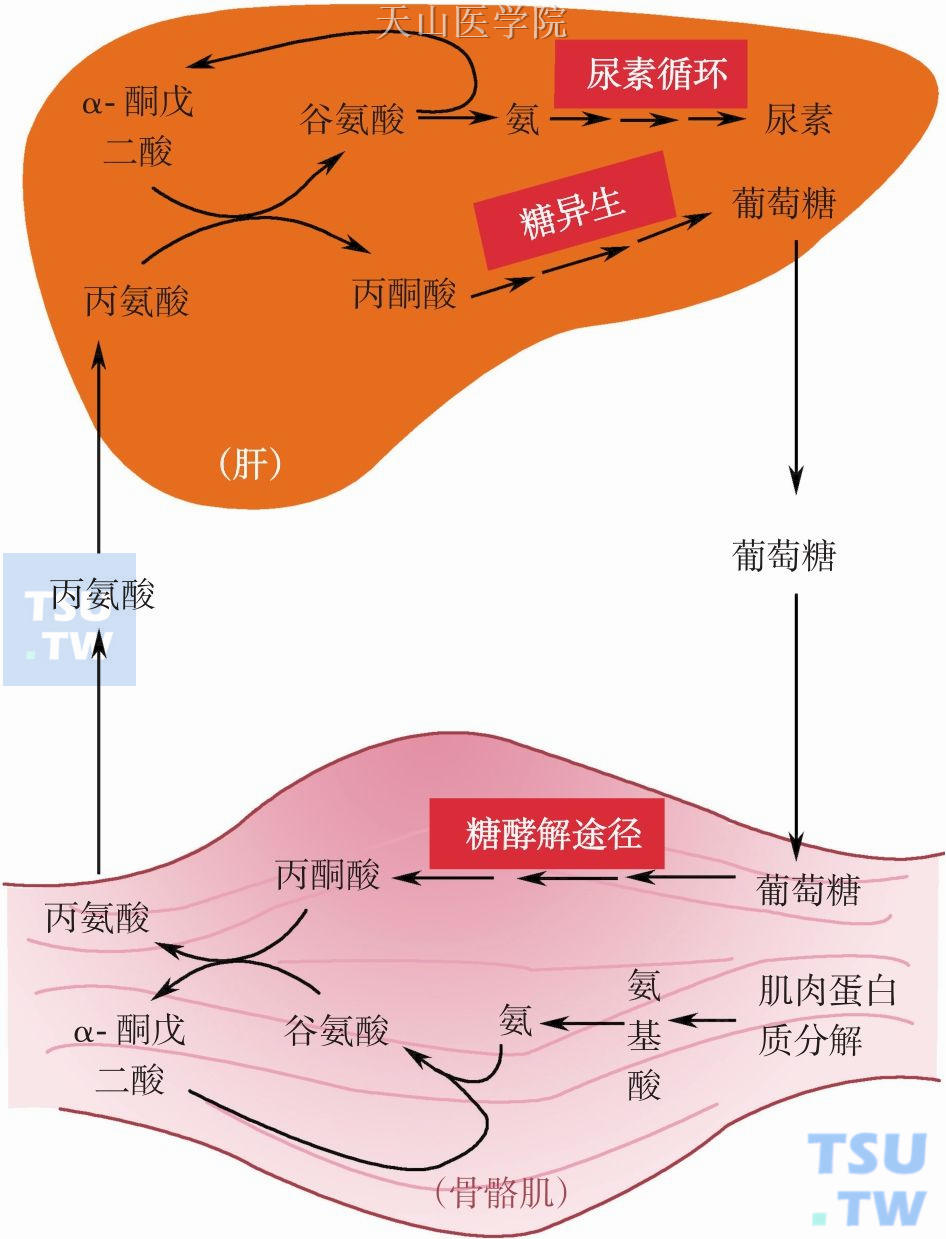
图3-1-8 骨骼肌与肝脏之间的丙氨酸-葡萄糖循环
