
人类对唾液腺及唾液的认识有悠久历史。J. R. Garrett等在其编撰的《唾液腺分泌的腺体机制》一书中对唾液腺分泌的机制做了详细的介绍。世界上有关唾液最早的记载是在公元前2000年左右的《亚叙植物志》中;其中描述了颠茄可抑制唾液分泌。有人认为更早的古埃及人亦有类似记载。早在第4世纪时,罗马皇帝的医师Orabasius(325~403年)就对唾液腺的基本结构与功能有清楚的阐述,其中对下颌下腺的描述如下:“在舌的两侧有导管的开口,将唾液排入口腔。这些导管源于舌根部的唾液腺。它们如同动脉一样,从腺体分支出来,导流唾液,从而湿润舌及相邻的口腔部分。”这种知识与现代唾液腺学十分接近。遗憾的是,这些科学知识很快就被世界遗忘了,代之而起的是迷信的臆测,即唾液腺是用来清除血液内的邪灵的。古人还认为唾液和胆汁、胃液及尿液一样,是一种体液,在体内循环流动。这种观点一直持续到文艺复兴时期(公元14~16世纪),极大地妨碍了对唾液腺的研究。直到显微镜发明之后,唾液腺和唾液的来源才重新被认识。
现已清楚,唾液腺和唾液有分泌、润滑、缓冲、清洁、凝集、排泄等功能,有帮助消化、协助味觉、维持口腔水分和抗微生物作用,可形成薄膜和离子池,并可用作诊断媒体。本篇先从唾液腺的分泌功能说起:
外分泌功能
唾液腺既有外分泌功能也有内分泌功能。迄今为止,绝大多数研究工作是以其外分泌功能为对象的。因此,我们关于唾液腺外分泌功能的知识远远多于对其内分泌功能的了解。
与大多数外分泌腺,如胰腺、乳腺、胃腺、前列腺、胆囊、汗腺等一样,唾液腺的主要外分泌功能是分泌该腺体所产生的特有液体——唾液。唾液的成分十分复杂,含有多种蛋白质和其他有机物、电解质和其他无机物及水分。
唾液的分泌量
关于唾液的分泌量,曾有人认为健康成年人每天分泌约1~1. 5L唾液,这个数值明显是一种过高的估计。人体混合唾液静止状态下流率为0. 3~0. 4ml/min,假设人的非睡眠期为15小时,所分泌的唾液应为270~360ml。睡眠时的最大分泌量为0. 1ml/min,如果睡眠时间为8小时,所分泌的唾液量应为48ml。据调查,人体用于进食的平均时间为每天54分钟,按60分钟计算;进食所刺激的流率为4ml/min,因而进食时所分泌的唾液量应为240ml。因此,人体每天唾液总分泌量应为558~648ml,远远低于许多书籍的记载。
在几组唾液腺中,分泌量最大的腺体是下颌下腺,占总唾液分泌量的60%~65%;其次为腮腺,占20% ~25%。舌下腺分泌量约占总唾液量的7%~8%,而小唾液腺分泌量较小,总共不超过7%~8%。
唾液流率
唾液流率也称为唾液分泌速率。大唾液腺的流率通常以ml/min或g/min表示。小唾液腺的排出口不易观察,其流率是用单位面积内每分钟收集的唾液容积来表示的,最常见的是μl/(cm2•min)。唾液流率是通过收集唾液来确定的,又分为非刺激流率(静止流率)及刺激流率。
唾液流率是衡量唾液分泌的最重要的指标。流率的减低反映了唾液腺功能的减低,表现为口干症,可明显影响口腔生理功能,例如语言、咀嚼、吞咽、味觉,也会损害口腔的防御机制,增加口腔疾病的发生率,如口腔黏膜感染、龋齿、牙菌斑形成、牙周病、细菌性唾液腺炎、口腔溃疡等等。
唾液腺的分泌取决于刺激状况。静止状态下,人体混合唾液的流率在0. 31~0. 39ml/min,平均为0. 32ml/min。但是,绝对静止状态是不存在的。所谓静止状态,仅仅是指没有明显的刺激因素而言。实际上,所有有关唾液流率的资料均来自于收集测定唾液量,而测定的操作过程本身即有刺激作用。例如测定时的张口和闭口,以及放置唾液收集器具等,均可刺激唾液分泌。然而,一般所说的刺激并不包括操作在内,而是指使用刺激物,如柠檬酸、糖类或受体激动剂。
静止状态下即在没有刺激时所分泌的唾液,主要来自下颌下腺和小唾液腺,这种分泌称为静止分泌或非刺激性分泌。非刺激性分泌常常包括自发性分泌,但是参与自发性分泌的腺体取决于动物的种类。羊的腮腺在静止状态下有大量分泌,而狗和猫的自发性分泌主要来自舌下腺。有人认为,大多数唾液腺均有自发性分泌,但有些则不能被观察到,因为分泌量很小,全部被导管系统重吸收了。这种看法缺乏理论基础和实验证据,理由是唾液腺导管系统并不重吸收水分。
早期认为小唾液腺只有持续的自发性分泌,但后来的研究发现,小唾液腺也有刺激分泌。小唾液腺的分泌速率远小于大唾液腺,因而一般用μl/(cm2•min)来表达。虽然有多种方法可用于测定小唾液腺的流率,但到目前为止,Periotron法被认为最准确,也是最常采用的方法。表3-1列出了小唾液腺的静止分泌速率。在小唾液腺之中,颊腺的分泌速率最高,唇腺次之,而腭腺较低。目前尚无舌前腺、舌后腺(von Ebner glands)、舌后侧腺(Weber glands)以及其他小唾液腺的测定资料。
当支配唾液腺的交感和副交感神经纤维受到刺激时,唾液可大量分泌,流率可增加许多倍。15~74岁的健康男性的非刺激流率平均为0. 36ml/min,用石蜡刺激时为1. 80ml/min;女性的流率稍低,非刺激流率平均为0. 26ml/min,刺激流率为1. 40ml/min。唾液大量分泌可见于进食咀嚼或味觉及嗅觉等神经受到刺激时。在各种唾液腺中,腮腺的刺激后分泌量增加最多,可达20倍以上。健康成年男性腮腺的平均非刺激流率为0. 08ml/min,女性为0. 07ml/min,用柠檬酸刺激后男女都增高为1. 62ml/min。
表3-1 人小唾液腺唾液的平均流率[μl/(cm2•min)]
![人小唾液腺唾液的平均流率[μl/(cm2•min)]](https://www.tsu.tw/kou/tuoyexian/img/image00701.jpg)
影响唾液流率的因素
(1)影响非刺激分泌的因素:
许多因素不同程度地改变非刺激唾液分泌,其中最重要的因素有:体水含量、光线、体位、生物节律、药物、性别、年龄以及心理因素等。
1)体水含量的影响:身体脱水时,唾液分泌减少;相反,水潴留时分泌增加。这也是日常生活中常常见到的状况,当大量出汗或蒸发时,人感到口干,缺乏唾液。假如失水量达8%,唾液分泌就会完全停止。
2)体位和光线的影响:站立位置可使唾液流率增高,而躺卧位可使流率减低。黑暗环境或遮光状态可使唾液流率减低30%~40%。但盲人的流率可能和正常人类似,这是长期适应的结果。
3)生物节律的影响:一日当中,唾液分泌速率以下午最高,睡眠时最低。这种节律变化也为睡前清洁口腔提供了依据。
4)药物的影响:许多药物可影响唾液分泌,其作用机制不同,有的是间接作用,有的是直接阻断唾液腺细胞膜上的受体。业已证明,可影响唾液分泌的药物有镇痛剂、食欲抑制剂、抗风湿药、抗胆碱能药物、止泻剂、止吐剂、抗组织胺药物、降血压药、利尿剂、治疗帕金森症的药物、抗焦虑剂、抗抑郁症药物、治疗精神病的药物。然而,并不是每种剂型均有抑制唾液分泌的作用。
5)性别和年龄的影响:唾液分泌速率有男性高于女性的趋势,机制不明。
成年人静止唾液流率与年龄无关,但老年人的分泌速率减低。早期的研究结果认为,多数老年人服用药物,分泌量减少的原因可能是药物的影响,从而认为唾液流率并无衰老性改变。但最近的调查表明,不服用任何药物的健康老年人的唾液流率明显低于年轻人。
6)心理的影响:看到食物及想象进食可能引起唾液的分泌,这就是所谓的心理-唾液腺反射,是一种条件反射。这种反射引起的分泌量增加不多。紧张、焦虑、兴奋、愤怒等情绪变化对唾液分泌有影响,但缺乏专门设计的研究。
7)疾病的影响:有些直接累及唾液腺功能的疾病如某些自身免疫性疾病,包括舍格伦综合征、类风湿病、红斑狼疮等可以引起唾液分泌的明显减少。头颈部恶性肿瘤放射治疗后亦可造成唾液分泌减少。这些疾病引起的唾液缺乏将在有关章节中专门介绍。
(2)影响刺激分泌的因素:
刺激唾液流率指的是唾液腺受到分泌性刺激时的分泌速率。最常见的刺激有进食和咀嚼。调查研究时所使用的刺激物有石蜡、糖块、柠檬汁及柠檬酸等。如上所述,刺激可使唾液分泌大量增加,但许多因素可影响刺激分泌速率。
1)刺激的性质:进食时的刺激是一种混合性刺激,它包含了咀嚼、味觉与嗅觉等多方面的刺激。单纯咀嚼运动所引起的唾液分泌增加小于咀嚼加味觉和(或)嗅觉的刺激。因而研究刺激分泌时,刺激的性质需要仔细设计。研究中常用的刺激物均可引起大量分泌,但增加程度变异较大。例如石蜡刺激时人混合唾液分泌速率为1. 6ml/min。咀嚼口香糖时的分泌速率为1. 7ml/min。葡萄味道的糖块所引起的腮腺流率为1. 0ml/min,而柠檬汁刺激为1. 5ml/min。因此,使用不同刺激所观察到的结果没有可比性。
2)味觉和嗅觉的刺激:四种味道刺激可明显增加唾液分泌,即酸、咸、苦、甜,其中以酸的刺激作用最强烈。研究表明,如用5%柠檬酸刺激,可获得最大的唾液流率,7ml/min。临床评价口干症时,常统一使用3%柠檬酸进行刺激。这样,所收集的结果则可进行不同方位的比较。嗅觉刺激的作用弱于味觉刺激,而且,嗅觉刺激也更难设计和评价。
3)吸烟的影响:吸烟对唾液分泌的影响评价不一。有些观察认为,吸烟或尼古丁可刺激分泌。由于控制唾液分泌的神经纤维含有烟碱(尼古丁)受体,血液中尼古丁含量的增加可以刺激分泌。唾液腺的腺泡细胞并无烟碱受体,因而尼古丁并不能直接作用于分泌细胞。也有一些研究认为,长期吸烟或使用尼古丁制剂有抑制分泌的作用,机制仍待阐明。
4)单侧咀嚼:有些人习惯在进食时用单侧咀嚼,这种不平衡的机械刺激可引起该侧的唾液分泌增加,但长期后果尚待进一步评价。
5)腺体大小:静止分泌的速率可能和腺体大小无关,但刺激分泌的流率与腺体大小成比例。
6)性别和年龄的影响:同静止分泌一样,刺激分泌流率呈男性高于女性的趋势。年龄对刺激分泌的速率有一定的影响。儿童的分泌速率较低,随年龄增高,到13~15岁时达成年人水平。5岁男孩和女孩的刺激流率分别为0. 67ml/min和0. 52ml/min,6岁时为0. 91ml/min和0. 77ml/min,8岁时为1. 17ml/min和0. 79ml/min,10岁时为1. 23ml/min和1. 01ml/min,13岁时为1. 90ml/min和1. 59ml/min,15岁时达2. 24ml/min和1. 92ml/min。老年人分泌速率减低(见第四节)。
7)进食的影响:进食是极为强烈的分泌刺激,但对这种作用研究甚少。这或许是由于进食刺激时,唾液难以收集。有人对7种食物的影响进行了研究。结果表明,进食水煮的大米只能引起相当于5%柠檬酸所刺激的分泌量的43%;而进食一种具有酸甜味的饼所刺激的分泌速率为最大速率的70%。进一步研究发现,食物的味觉刺激的作用远远强于单纯咀嚼的刺激。咀嚼口香糖即是一个明显的例子。口香糖在最初10分钟内可引起很高的唾液流率,其后随着口香糖的味道的减弱,唾液分泌速率减低。
8)药物的影响:同静止分泌一样,刺激分泌明显受药物的影响。临床常用的药物大多数对唾液分泌有抑制作用,例如利尿剂、皮质激素、镇痛剂和三环抗抑郁剂、β受体阻滞剂、降血压药、α甲基多巴、治疗糖尿病的药物、可乐定等均可使唾液分泌减少。有些药物可减低唾液流率35%以上。只有硝酸盐可使唾液流率增加20%左右。许多老年人需服用不同种类的药物,药物的联合使用可能会加重对唾液分泌抑制。发展纠正这种副作用的新方法,是一个重要的研究领域。
内分泌功能
唾液腺合成并分泌入血的肽类物质达30种以上。这种分泌功能存在于许多动物种类,在啮齿类动物(小鼠、大鼠和地鼠)最常观察到。常见的分泌入血的激素样物质或调节肽有:表皮生长因子、神经生长因子、腮腺激素(parotin)、高血糖素、促胃液素、紧张素、促红细胞生成素、心房肽(心钠素)、大活动蛋白、S-100蛋白、神经元特异烯醇化酶、抑生长素、突触泡蛋白、嗜铬粒蛋白、降钙素等。
虽然不同的唾液腺合成、储存、分泌内分泌多肽,但绝大多数分泌入血的肽类是由下颌下腺合成并分泌的,舌下腺可能也有类似的功能。只有少数动物的腮腺导管系统中可见到某些肽类,如猪的腮腺可合成高血糖素和促胃液素,而狗的腮腺可合成促胃液素。腮腺素为腮腺所分泌。
唾液腺内分泌物质的合成几乎全部是在导管系统。导管系统的颗粒导管(GCT)细胞及纹管细胞是合成并储存这些多肽物质的主要场所(表3-2)。由于啮齿类动物有发达的颗粒导管系统,所以常用来研究这些多肽物质的合成与分泌。人类唾液腺没有颗粒导管系统,内分泌功能可能是在于纹管细胞。
各种多肽在细胞内的存储形式也大不相同。表皮生长因子和神经生长因子储存于分泌颗粒内,而其他调节肽则散在于细胞质内,只有胰岛素样蛋白聚集在分泌颗粒周围,机制不明。
这些多肽的分泌机制仍不清楚。胰岛素样蛋白对细胞外葡萄糖浓度有反应,当葡萄糖浓度增高时分泌。血浆葡萄糖浓度减低和精氨酸浓度增高可引起大鼠下颌下腺分泌高血糖素。迄今为止,很少观察到除生长因子以外的多肽被分泌到唾液中,因而有人认为,所有唾液腺细胞内的调节肽的释放都是内分泌过程。有人提出,激素样物质进入血液的途径可能有三种:①直接由导管细胞从基侧膜分泌到血液中;②先分泌到初始唾液中,然后由导管细胞重吸收入血;③先分泌到唾液中,在胃肠道中吸收入血。实验发现,从唾液腺流出的静脉血中含有血管舒缓素和紧张素,证明这些物质是由唾液腺直接分泌的,而不是由胃肠道吸收入血的。已知颗粒小管细胞的基侧膜可以进行出胞作用。此外,尽管导管细胞可以重吸收蛋白质,但尚无证据表明重吸收的蛋白质可以进入血流。如果多肽物质被首先分泌到唾液中,在胃肠道中不被酶分解而以原形被吸收的机会极小。因此,唾液腺内分泌功能不可能通过先分泌到唾液中的过程而实现。
表3-2 啮齿类动物下颌下腺多肽物质的细胞分布
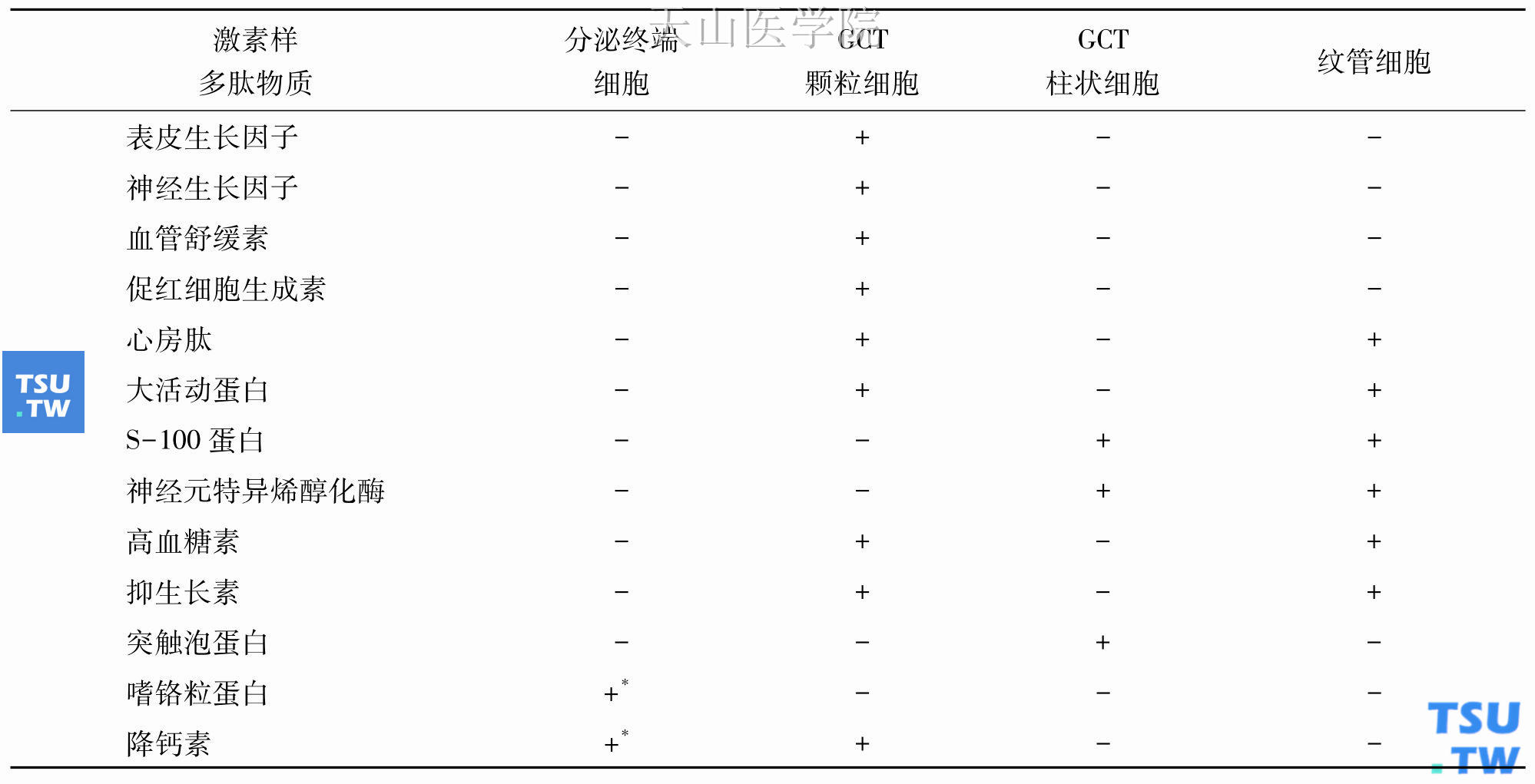
*仅见于生后早期的细胞中。
腮腺激素
腮腺激素最初是从猪腮腺组织中分离出来的,是一种富含脯氨酸和甘氨酸的小分子糖蛋白,分子量为8100。腮腺激素的作用是调节蛋白质合成、促进软骨钙化及减低血钙。腮腺激素也可调节牙组织内液体的流动过程。当给大鼠注射荧光染料时,腮腺激素可促进荧光染料从牙髓中穿过牙本质而达到釉质牙本质界。
腮腺激素分泌的调节尚不清楚。一种看法认为,腮腺激素可能是下丘脑-腮腺内分泌轴的一个重要组成部分。下丘脑分泌一种释放因子,使腮腺分泌腮腺激素到血液中,引起牙本质内染料的转运。腮腺激素释放因子已从猪下丘脑-丘脑组织中分离出来,当静脉注射到大鼠和猪体内时,这种因子可引起腮腺激素的分泌。此外,已知摄食是一种引起腮腺激素分泌的强刺激。狗在摄食后的1分钟内血液中即可出现腮腺激素的增高,10~20分钟时达高峰,以后逐渐减低,但可持续3~4小时之久。大鼠摄食时,牙本质内的染料穿透增强,但高糖食物可使大鼠牙本质内的染料转移减少。猪摄入高糖食物时分泌腮腺激素水平低于摄入常糖食物。
人腮腺是否有类似激素,尚不清楚。
生长因子
唾液腺导管系统的颗粒导管上皮可合成生长因子,包括表皮生长因子、神经生长因子及其他生长因子。这些生长因子通常储存在分泌颗粒内。刺激α-肾上腺素能受体可使这些因子释放到唾液中。刺激胆碱能受体也可引起类似的释放,但作用弱于α-肾上腺素能受体。这些生长因子的分泌入血过程及调节仍不清楚。
(1)表皮生长因子(EGF):
Turkington等(1971)首次发现EGF存在于小鼠下颌下腺的颗粒小管(GCT)细胞内,主要储存部位是分泌颗粒。后来用原位杂交技术证明,EGF mRNA在GCT细胞中大量表达。EGF也大量存在于大鼠和地鼠的下颌下腺GCT细胞中。事实上,成年大鼠体内的EGF主要是由肾和唾液腺合成的。EGF的量在雄性动物明显高于雌性。而且,用雄性激素处理动物可使EGF及其mRNA大量表达。切除雄性动物睾丸后,EGF减低到雌性动物水平。孕激素可能有类似作用,但尚有争议。给切除睾丸后的雄鼠及正常雌鼠注射孕激素可引起EGF含量的增加。而且,妊娠和哺乳的动物GCT细胞内EGF水平明显增高。与之相反,雌激素抑制EGF及其mRNA的表达,切除卵巢后表达增强。切除肾上腺的小鼠,细胞内EGF的表达大为减少。给予肾上腺皮质激素如可的松、地塞米松可使EGF表达恢复。甲状腺激素也可使EGF及其mRNA表达增加。与之相符,甲状腺功能低下时EGF水平明显减低。
Jones等(1995)观察到,EGF是肝再生必需的因子。切除大鼠肝的三分之一,12天后,肝可恢复到正常重量的89%。如果同时切除大唾液腺,12天后肝的重量仅为正常的55%。给这些大鼠注射外源性EGF可使肝的再生能力恢复。EGF的作用是刺激肝细胞合成DNA。这些结果表明唾液腺EGF分泌入血具有重要的生理作用。
EGF的合成与发育相关。小鼠生后2周内唾液腺中并不能检出EGF,到第20天时则可检出。精确地讲,EGF蛋白质和mRNA可在生后第18~20天时测出。
(2)神经生长因子(NGF):
免疫组化研究发现,NGF存在于小鼠与大鼠的下颌下腺GCT细胞中。原位杂交技术证明,NGF mRNA确实是在GCT细胞内表达。NGF的主要储存部位是分泌颗粒,表明其分泌过程可能为胞吐作用。研究证明,刺激蛋白质分泌以后,细胞内的NGF水平减低。Zn2+可以稳定NGF的结构。研究发现Zn2+与NGF共存于分泌颗粒内。
细胞内NGF水平受雌激素调节。甲状腺激素亦可使NGF mRNA的表达增加。另外,给予成年小鼠可的松龙可增高细胞内NGF水平,表明肾上腺皮质激素亦参与NGF表达的调节。最近的研究发现,褪黑激素可能也参与NGF合成的调节过程。NGF水平与GCT系统发育平行。小鼠生后3周时可检出NGF。在3~12周之间,NGF mRNA表达逐渐增加。
(3)生长因子的分泌:
EGF和NGF可被大量分泌到唾液中,但这些生长因子在唾液中的精确功能仍不清楚,可能与促进口腔及胃黏膜的生长发育和修复有关。根据这种假设,有人曾用生长因子来治疗胃溃疡,获得良好效果。
生长因子分泌入血的过程尚不明了。切除小鼠下颌下腺后并未观察到血浆内生长因子的改变,从而对其分泌功能产生怀疑。但是血液内通常不易检出生长因子,因而上述实验的推论还有待于进一步证实。实验研究发现,刺激α-肾上腺素能受体或者引起动物争斗行为可促使生长因子释放入唾液,同时血浆水平也大大增高。这些结果提示,应激反应也许是激发生长因子分泌入血的重要因素。然而,有人证明,切除肾上腺皮质以及阻断唾液腺的肾上腺素能受体并不影响NGF的分泌入血。
血管舒缓素(kallikrein)
人体唾液腺可合成一种激素样物质血管舒缓素。该物质可分解激肽原(kininogen)形成激肽(kinin),因而又称为激肽释放酶。激肽可使平滑肌松弛,从而使血压减低。Ellison 等(1967)发现,刺激鼓索神经可引起下颌下腺血管扩张,这种作用来自于血管舒缓素。
自1936年Werle和Von Roden报道血管舒缓素存在于唾液腺中以来,唾液腺已被公认是可合成此酶的器官。下颌下腺的GCT细胞是该酶的储存场所。腮腺和舌下腺血管舒缓素的含量很小。血管舒缓素的活性结构可能是rK2~rK10,rK1的生理功能仍不清楚。已有证据表明大鼠下颌下腺血管舒缓素可释放入血,因为切除唾液腺后的24小时内血中血管舒缓素浓度减低为36%。热应激可引起血管舒缓素分泌入血。电刺激交感神经可引起大量血管舒缓素释放入血。例如,用10Hz刺激1分钟即可引起这种效应,并伴随激肽浓度的增高。以50Hz刺激单侧交感神经1秒钟,间隔10秒钟,共60分钟,可使血清rK1浓度增加48%。与之类似,刺激副交感神经使血清rK1浓度增加46%。切除下颌下腺和舌下腺后4天,血清rK1浓度减低50%。这些研究清楚表明,生理状态下下颌下腺分泌rK1到血清,而且这种分泌与分泌到唾液中的量无关。血清中存在有血管舒缓素抑制蛋白,可使入血的血管舒缓素很快失去活性。
肾素
肾素存在于小鼠的下颌下腺中,雄性高于雌性。雄激素对其有调节作用。雄性小鼠生后25天时即可在GCT细胞中检出肾素,雌性小鼠要到30天时才可检出,晚于EGF和NGF的检出时间。精细的免疫组化分析证明,肾素存在于GCT细胞的分泌颗粒及致密囊泡中。此外,肾素的表达有很大的个体细胞差异。大多数GCT细胞表达EGF和肾素,但有些GCT细胞只表达EGF而不表达肾素。对此,Bing等(1981)认为这是固定样品时处理不当所致。但近年来研究表明,肾素的合成比较复杂,可形成多种蛋白质形式,例如单链和双链蛋白质等等。
肾素的蛋白加工过程受雄激素的调节,表达肾素受遗传的控制。有些种类的小鼠表达高水平肾素,有些种类则表达低水平。这种遗传变异是由于小鼠中含有两种结构不同的肾素基因Ren-1和Ren-2。含有Ren-1基因的小鼠GCT细胞中肾素很低,而Ren-2的基因可使GCT细胞合成大量肾素。Ren-2基因也显示出发育和激素调节的组织特异性。给仅含有Ren-1基因的小鼠受精卵表达Ren-2基因,从而产生转基因小鼠,这种小鼠下颌下腺合成高浓度的肾素,其合成过程受睾酮和甲状腺素的调节。大鼠的GCT细胞不能合成肾素,即使把Ren-2基因转到大鼠细胞内,仍无肾素合成。
促红细胞生成素
唾液腺被认为是促红细胞生成素合成和分泌的后备器官。小鼠下颌下腺中可合成促红细胞生成素。然而,正常小鼠唾液腺中并不能检出该物质。在贫血、缺氧及肾切除后则可检出,说明其合成受红细胞功能状态的调节。此外,促红细胞生成素的合成和分泌可能受性激素的调节。用睾酮处理小鼠可使唾液腺颗粒导管细胞合成促红细胞生成素大量增加。
心房肽
心房肽又名心钠素,是一种激素类物质,是由心脏细胞合成分泌的,参与血压和体液的调节。近年的研究发现,唾液腺中也含有高浓度的心房肽,主要存在于颗粒导管细胞和柱状细胞内。目前对心房肽的分泌过程知之甚少。现已观察到,动物在缺水状态下,心脏细胞中心房肽含量尚未发生改变,唾液腺颗粒导管细胞中心房肽已经减低,说明唾液腺心房肽的调节更为灵敏,也提示唾液腺内的心房肽可以分泌。目前对唾液腺心房肽在全身性血压和体液的调节过程中的作用几无了解。
