
凝血活化的外源途径在生理止血中起作用。受损血管壁释出的组织因子(TF),在Ca2+参与下,促使血浆FⅦ活化成Ⅶa而启动凝血。FⅦa- TF复合物具有双重作用:即能锚钉在细胞膜上使凝血局限于受损部位,又能直接使FⅩ和Ⅸ(内源因子)活化。外源凝血系率先生成少量FⅨa、Ⅹ、启动性凝血酶和纤维蛋白。启动性凝血酶不仅能使邻近的血小板活化并通过正反馈机制激活坐落在血小板磷脂表面的FⅤ(FⅩ受体)、FⅧ,在活化血小板表面来加速内源因子的活化。正反馈机制能形成高速率的凝血酶原酶,使FⅩa得到补充和加强,来达到最终止血。实验室中常采用凝血酶原时间(prothrombin time,PT)试验作为外源凝血系的监测试验。
组织因子的作用
组织因子(tissue factor,TF)是一种跨膜糖蛋白,Mr 53 000,由脱辅基蛋白和磷脂组成。TF广泛存在于大多数非血管细胞表面及血管外膜层,是多种细胞(平滑肌细胞、成纤维细胞)的膜内蛋白,也存在于脑(灰质甚于白质)微血管周围。培养的内皮细胞或平滑肌细胞在各种激动剂的刺激下表达TF的同时尚伴有细胞内Ca2+动员及蛋白激酶C的反应。TF可分成非血管细胞的结构性TF及血细胞(主要是单核细胞)和内皮细胞的可诱生性TF。TF是FⅦ和FⅦa的膜受体,是两者的辅助因子,能使Ⅶa的催化功能提高1000倍。但是正常情况下血液中不出现TF。当血管受损或撕裂,或内皮细胞、单核细胞受内毒素、补体C5a、免疫复合物、IL- 1、TNF等介质攻击时,结构性或诱生性TF即与血流接触,立即形成细胞表面的FⅦa- TF复合物。这一复合物的形成即启动组织因子途径终至生成凝血酶及形成纤维蛋白凝块。
FⅦ的活化
FⅦ属于单链糖蛋白,Mr 5000,血浆水平为10nmol/L。FⅦ及TF本身无促凝活性。FⅦ内源(水解)酶的表达必须存在辅因子TF,两者组成复合物后的TF能使FⅦ自我活化成Ⅶa。FⅦa- TF中酶原性Ⅶ单一多肽链的精氨酸152-异亮氨酸153键裂解,产生的Ⅶa由轻链(Mr 20 000)和重链(Mr 30 000)以二硫键相连(图1)。TF膜内锚钉区缺失即丧失其使FⅦ自我活化能力,但是仍保留其辅因子(增强Ⅶa催化)功能。FⅦa能激活它的两个底物Ⅹ和Ⅸ。
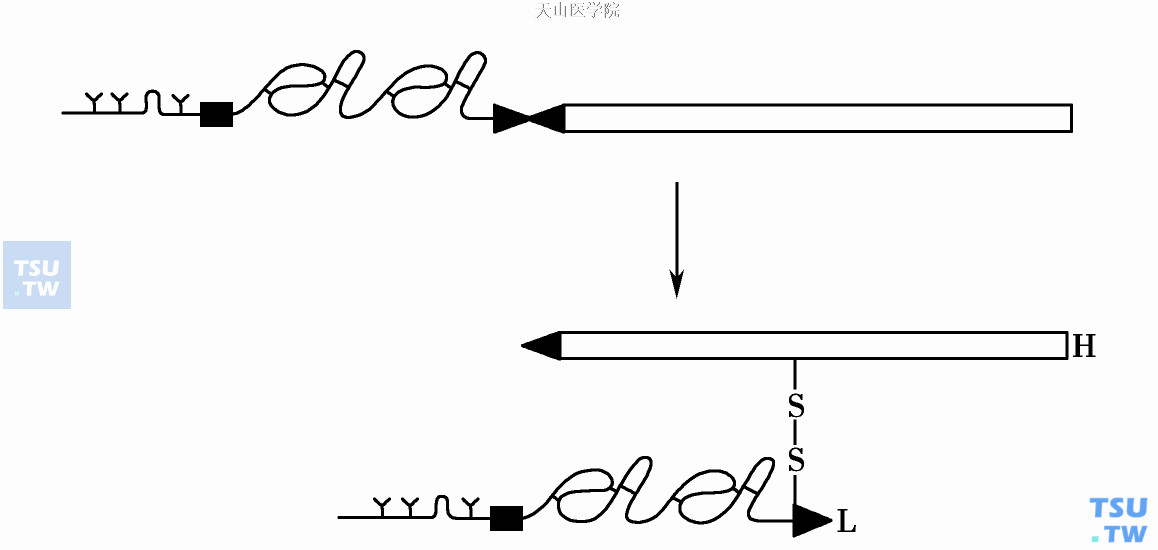
图1 FⅦ的活化过程(L轻链,H重链)
FⅨ的活化
外源系FⅦa- TF复合物及内源系FⅪa能活化FⅨ。两条途径活化动力学过程不同,但有着相同的裂解部位和产物。和其他维生素K依赖性凝血因子的活化方式相同,FⅨ活化必须在血液中存在Ca2+,反应必须在膜表面(血小板或人工磷脂小粒)发生;单纯的膜表面不能加速FⅨa生成。
单链型FⅨ酶原蛋白活化成Ⅸa(αβFⅨ),通过二次裂解:第一部位精氨酸145-丙氨酸146,之后第二部位精氨酸180-缬氨酸181。FⅨa有轻链(L)Mr 18 000(氨基酸1~145)和重链(H)Mr 27 000(氨基酸181~416)以二硫键相连(图2),活化过程中释出活化肽片(activation peptide,AP)名FⅨP(氨基酸146~180)。FⅨa及FⅨ通过Ca2+或其他金属离子桥连于磷脂膜表面。FⅨa含暴露的Ⅷa辅因子结合位点及酶催化区,能进一步结合Ⅷa并使FⅩ活化。
虽然接触激活系的FⅪa(内源因子)也能激活Ⅸ,例如PTT测试中Ⅸ的体外活化,但是缺乏Ⅺ(丙型血友病)者很少有出血表现,故推测Ⅺa激活Ⅸ并非唯一的FⅨ活化途径。FⅩa也能激活Ⅸ使后者两个部位裂解,但生成速率十分缓慢,似乎不能参与正常止血。推测生理情况下,FⅨ活化是通过混杂(Ⅶa- TF及FⅩa)机制,使第一部位裂解产生中间产物Ⅸα,之后接着由Ⅶa- TF使中间产物活化。纯系统体外实验所得的蛋白超过生理状态下获得的量支持上述推测。中间产物可能与增强活化速率有关,但其功能尚不完全清楚。
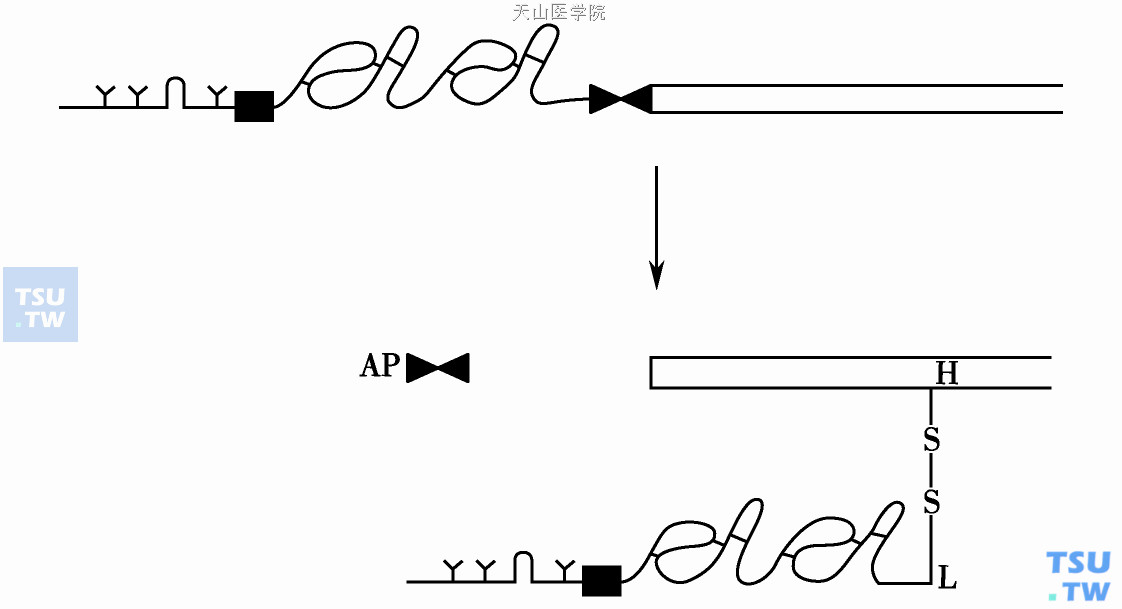
图2 FⅨ的活化过程;FⅨa由轻链(L)和重链(H)以二硫键相连,活化过程中释出活化肽片(AP)
FⅩ的活化
FⅩ又名Stuart- Prower因子,是内外凝血系汇合处共同通路中的凝血因子,可被FⅨa-Ⅷa复合物(Ⅹ酶,曾名内源Ⅹ酶)或FⅦa- TF复合物(曾名外源Ⅹ酶)所激活。Ⅹ酶复合物中FⅧa为辅因子。无活性的单链FⅧ几乎不具有辅因子活性。
血循环中的FⅧ以Ca2+依赖方式和大分子vWF结成二聚体。FⅧ的重链(Mr自90 000~200 000)和轻链(Mr 76 000)以非共价键结合。vWF能抑制FⅧ轻链和血小板膜微粒以及阴性磷脂的结合。FⅧ是凝血酶或Ⅹa的底物,被后二者活化。FⅧ(功能区自N端起A1- A2- B- A3- C1- C2其前三区为重链,后三区为轻链)被凝血酶或Ⅹa反馈性裂解至少2~3个键(A2- B接连点- B链切除,A1- A2分离及释出vWF分子),其中精372- 373,精1686- 1689的裂解是FⅧ活化所必需。FⅧa(异三聚体)由来自重链A1区(Mr 50 000)、来自A2区带着部分轻链(Mr 43 000)和来自轻链A3C1C2区(Mr 73 000)的三种亚单位以非共价键结合而成。FⅧ和Ⅷa都能与活化血小板表面Ⅷa受体区(阴性磷脂)紧密结合。凝血酶活化或Ⅹa活化的Ⅷa特性不同,分别以Ⅷa(Ⅱa),Ⅷa(Ⅹa)表示;Ⅷa(Ⅹa)①仅具有30%~40%Ⅷa(Ⅱa)辅因子活性;②能使FⅧa(Ⅱa)的催化速率衰退40%;③可被凝血酶(Ⅱa)转化成更活化的形式FⅧa(Ⅱa)。
FⅨa和Ⅷa通过受体结合在磷脂活化血小板表面必需Ca2+的方式组成(Ⅹ酶)复合物。内源Ⅹ酶激活FⅩ存在着生理时相过程,因为在血小板活化后4分钟内FⅧ受体在膜表面才达峰值。启动凝血酶产生大量活化血小板的正反馈机制能加速这一过程。同样,膜表面组成FⅦ- TF(外源Ⅹ酶)复合物也需Ca2+,以及外源Ⅹ酶活化FⅩ也存在着生理时相。
FⅩ是凝血系中唯一的双链酶原蛋白,Mr 59 000有重链(H)和轻链(L)以二硫键相连。血浆FⅩ水平约170nmol/L。FⅩ重链精51-异亮52,单一的键裂解(图3),释出高度糖基化的活化肽段FXP Mr 7000,并产生FⅩa。酶原蛋白FⅩ活化只引起结构细微改变,但却发生重要的蛋白酶功能。和所有丝氨酸蛋白酶相同,FⅩa酶活性的表达涉及重链的组氨酸93-天冬氨酸138-丝氨酸233位三合一的组装反应。FⅩ能被Ⅹa反馈性活化(裂解苏氨酸位),但过程很慢,不参与生理止血。
鲁塞尔蝰蛇毒时间(Rusell’s viper venom time,RVVT)(又名:stypVen时间)测定,是利用RVV凝血蛋白能活化FⅩ以及其类凝血酶特性加强Ⅴ促凝作用,使富血小板血浆凝固,凝固过程不需Ⅶ。在RVV作用下FⅩ活化肽(FⅩP)与Ⅹa仍相连,且Ⅹ的C端不参与Ⅹ酶复合物形成过程。RVVT临床应用于FⅩ缺乏者RVVT延长与Ⅶ缺乏者RVVT正常,相鉴别。
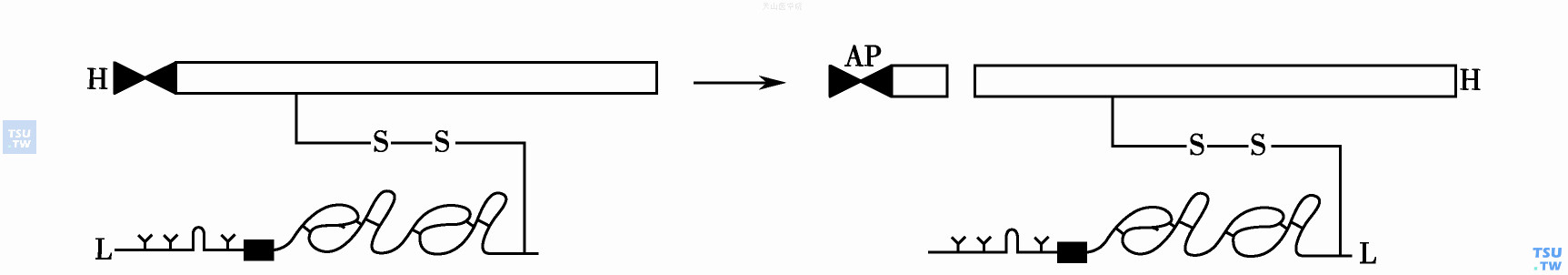
图3 FⅩ的活化过程
FⅩ酶原蛋白由重链(H)和轻链(L)以二硫键相连,FⅩ重链单一的键裂解释出活化肽片AP并产生FⅩa
FⅩa的生理性蛋白酶抑制剂包括抗凝酶Ⅲ(ATⅢ)、α2巨球蛋白和组织因子途径抑制物(tissue factor pathway inhibitor,TFPI)。ATⅢ和α2巨球蛋白能清扫及中和流经组织损伤部位血流内的FⅩa促凝活性。TFPI的生理功能最近才被充分认识。TFPI基因位于2号染色体,含9个外显子。TFPI Mr 34 000含有三个前后串联的kunitz型蛋白酶抑制区。TFPI能直接和FⅩa结合并形成FⅩa-TFPI-Ⅶa- TF抑制复合物,从而抑制Ⅶa- TF活性。抑制复合物通过下调TF促凝活性、迫使持续止血所需的Ⅹa必须通过内源Ⅹ酶来提供。
凝血酶原的活化
凝血酶原(prothrombin,PT)作为凝血酶原酶(FⅩa-Ⅴa)的底物而活化。凝血酶原酶(prothrombinase)是FⅩa和辅因子(FⅤa)在膜表面及Ca2+存在下组装成的复合物。
FⅩa-Ⅴa复合物(凝血酶原酶)的组装
凝血酶原酶组装图(图4)中三种维生素K依赖凝血因子(FⅩ、FⅤ、FⅠ)的γ- Gla区(以- Y代表),与Ca2+(以- O代表)相互反应。钙的占据使这些蛋白暴露出膜结合位点C(以黑点代表脂质结合位点)。辅因子FⅤa是FⅩa的膜受体,是凝血酶原活化反应中绝对依赖的辅因子。对凝血活酶功能起调节作用。
1. FⅤa作为FⅩa膜受体促进凝血酶原酶生成FⅤ和Ⅴa是膜外结合蛋白,能以高亲和力结合血小板膜表面的丝氨酸磷脂和活化血小板衍生的微粒,这一结合不需金属离子或Ca2+的参与。凝血酶或FⅩa将无活性Ⅴ转变成Ⅴa(辅因子),通过三处肽键裂解:精709-丝710、精1018-苏1019、精1545-丝1546。所产生的FⅤa含重链(Mr 110 000)和轻链(Mr 78 000)在Ca2+存在下非共价连接。FⅤa与血小板表面特殊部位结合即能作为FⅩa膜受体而确定凝血酶原酶的形成。
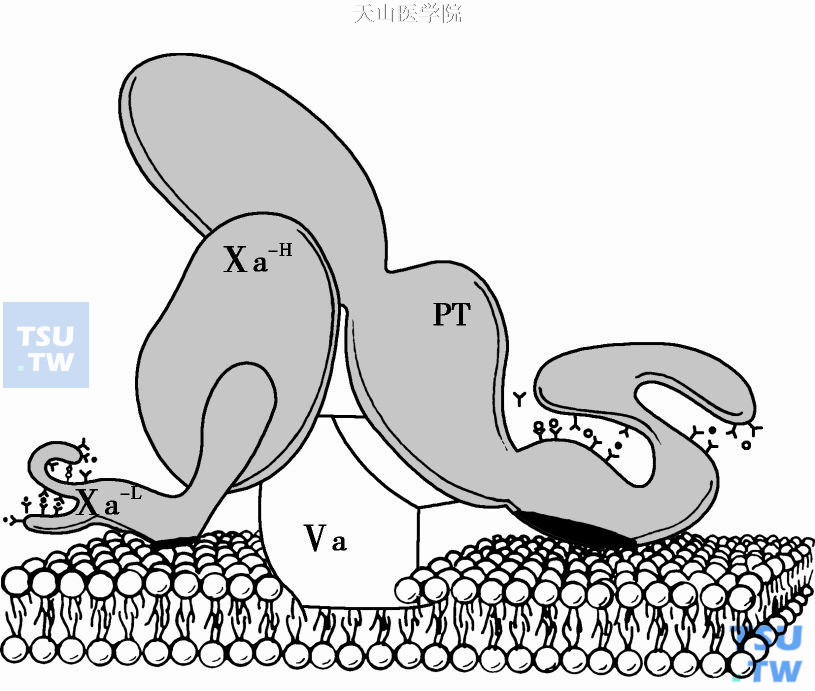
图4 凝血酶原酶组装图
2. FⅤa的辅因子功能 常采用血小板表面作为磷脂微粒模式来研究凝血酶原酶的组装过程,其细节可有多种变化。Ca2+存在下,膜表面的FⅩa即显示有蛋白水解酶的活性,直接水解并活化凝血酶原,但反应速率较凝血酶原酶慢300 000倍,不符合生理止血时相。活化血小板表面结合着的FⅤa以轻链与Ⅹa EGF、催化区结合,重链与凝血酶原结合。故膜结合着的FⅩa、FⅤa可通过相互反应所发生的构型改变使酶(复合物中的Ⅹa)与底物(凝血酶原)更接近。此外,FⅤa能与凝血酶原Kringle- 2反应以及由膜磷脂参与使Ⅹa活化凝血酶原的催化速率增加105数量级。FⅤa甚至能和内皮、单核、淋巴细胞膜结合而促使凝血酶原酶生成。
凝血酶的生成
单链的凝血酶原被FⅩa-Ⅴa(凝血酶原酶)复合物经2~3次肽键裂解后释出多种肽段并生成双键的凝血酶。凝血酶Mr 38 000,A- B两链由二硫键相连(图5)。
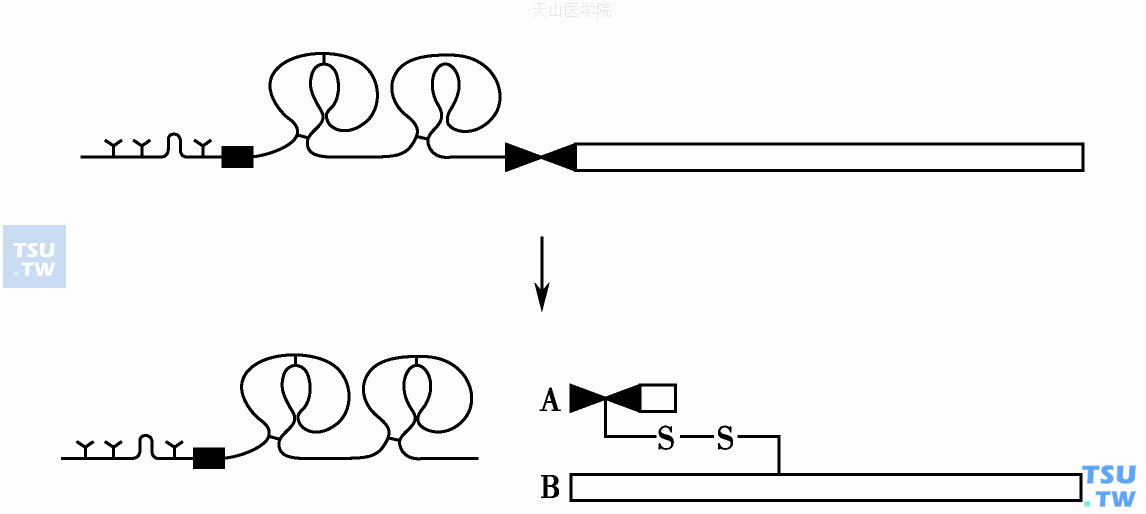
图5 凝血酶原的活化
凝血酶原活化需裂解2~3处肽键后释出多种肽段并生成双链的凝血酶,A- B两键由二硫键相连
凝血酶原分子至少有两种裂解方式:精氨酸271-苏氨酸272,或精氨酸286-苏氨酸287裂解。分别自N端释出活化的肽段F1+F2肽(Mr 43 000)或F1+F2+F3肽(Mr 45 000),以及产生相应的中间产物前凝血酶原2(prethrombin 2,Pre- 2)Mr 38 000或是N端延伸13个氨基酸的前凝血酶原2(meizothrombin)。Pre- 2无蛋白酶活性,需随即被FⅩa进一步水解才变成凝血酶。即Pre- 2及meizothrombin的C端精氨酸322-异亮氨酸323裂解,两者都能生成凝血酶。生成的凝血酶A链(Mr 5000)和B链(Mr 32 000)以单一的二硫键相连。Meizothrombin只能水解较小的底物,不能将纤维蛋白原转化成纤维蛋白。凝血酶的丝氨酸蛋白酶活化中心位于组氨酸365、天冬氨酸419、丝氨酸527。
凝血酶的促凝活性能被血浆ATⅢ所中和,肝素能增强ATⅢ的这一中和速率。
纤维蛋白原转化成纤维蛋白
纤维蛋白原(α2β2γ2)是血浆中含量最大的凝血因子,以非活化的形式作为血浆蛋白而循环。在凝血的最后阶段,纤维蛋白原被凝血最终生成的酶-α凝血酶裂解其Aα和Bβ链,转化成纤维蛋白(图6)。
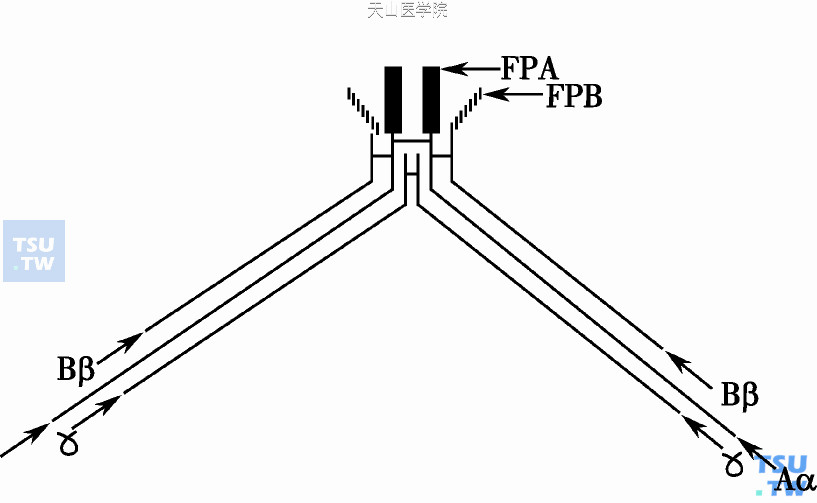
图6 纤维蛋白单体的生成;纤维蛋白原(α2、β2、γ2异二聚体)裂解后释出FPA、FPB并生成纤维蛋白单体
凝血酶和纤维蛋白的中央区(E)结合,裂解Aα 链N端精氨酸16-甘氨酸17键,释出2个各含16个氨基酸的纤维蛋白肽A(FPA)、Aα链产生新N端;又从Bβ链N端裂解精氨酸1-甘氨酸15键使释出2个各含14个氨基酸的纤维蛋白肽B(FPB)、Bβ链产生新N端。纤维蛋白原释出一对FPA和一对FPB后,转化成纤维蛋白单体。纤维蛋白丝的延长和加粗是通过单体的同种多聚作用:经肽键裂解所产生的单体暴露出中央区(E)的互补结合位点,能和其他单体C端(γ链的D区)非共价E- D互补结合而形成两股半重叠延伸的原纤维丝。含E- D连接区的原纤维丝再与邻近的第三个单体γ链D区通过D- D尾结合而聚集成粗纤维。非共价结合的原纤维多聚体是不牢固的,必须通过FXⅢa的作用使邻近的纤维蛋白单体分子间γ或α链共价交联而形成稳定的纤维蛋白多聚体。FXⅢa是一种Ca2+依赖性转谷氨酰胺酶,催化赖氨酸的ξ氨基和谷氨酸的γ-酰胺基间形成等肽键;主要连接部位:①纤维蛋白γ链C端形成γ-γ二聚体使分子加强并纵向增长;②连接α链延长的C端间,使纤维蛋白多聚体有纵及横向连接的两种加强方式。FXⅢ是酶原蛋白,需经过足量凝血酶的蛋白溶解作用,暴露出巯基活化中心而活化成XⅢa。纤维蛋白(原)的早期降解产物能加速XⅢa的活化,XⅢa尚可作为凝血酶敏感蛋白和纤维连接蛋白的锚钉蛋白、抛锚寄托场所。
