
1912年Mailard证实氨基酸可与还原糖形成稳定的酮胺化合物(ketoamine),该反应亦称Mailard反应。此后又发现这种反应广泛的存在于奶类及某些食品中,由于其反应物可变成棕色故又称Browning反应。当牛奶成食物加热时,糖中的醛基可与蛋白质中的氨基结合形成醛亚胺(aldimine),即Schiff碱,这是一个不稳定的中间产物,可还原回原来的反应物状态,它也能再发生一个分子中的结构重排,变成稳定的酮胺化合物被称为Amadori重排。重排反应是不可逆的,因此重排后的产物是稳定的。酮胺化合物中的糖部分,另外还有一个稳定化的趋势,即形成五环糖(furanose)或六环糖(pyranose)结构,它们是不需要酶催化的非酶促糖化反应,该反应符合质量作用定律,即反应率决定于反应物的浓度,当糖的浓度增高时,糖化反应产物便增多,反之,便减少。
血红蛋白同样具有与通过红细胞膜的葡萄糖进行非酶促糖化反应的特性。这种反应是在红细胞自然寿命的120天中在HbA生成后不断地进行着,这便是Bunn所提出的HbA1c生物合成学说,即HbA中每一条β链的N端缬氨酸的氨基可与1分子葡萄糖中的醛基发生反应而生成Schiff碱,这种不稳定的醛胺结构又自动发生Amadori重排反应而生成稳定的酮胺结构,后者是一种1-氨基-1-去氧-2酮葡萄糖的衍生物即HbA1c。HbA1c的形成并非由于基因的变化而产生,而是在体内首先合成HbA,在以后的代谢过程中又与葡萄糖加合而形成,所以HbA1c的形成过程称为合成后变化或转译后变化。又因为在血循环中的红细胞没有合成新蛋白质的功能,因而进一步说明HbA1c是HbA合成后经化学修饰而形成的。
HbA1c的生物合成过程根据其反应速度快慢的不同可分为两个阶段(下图)。
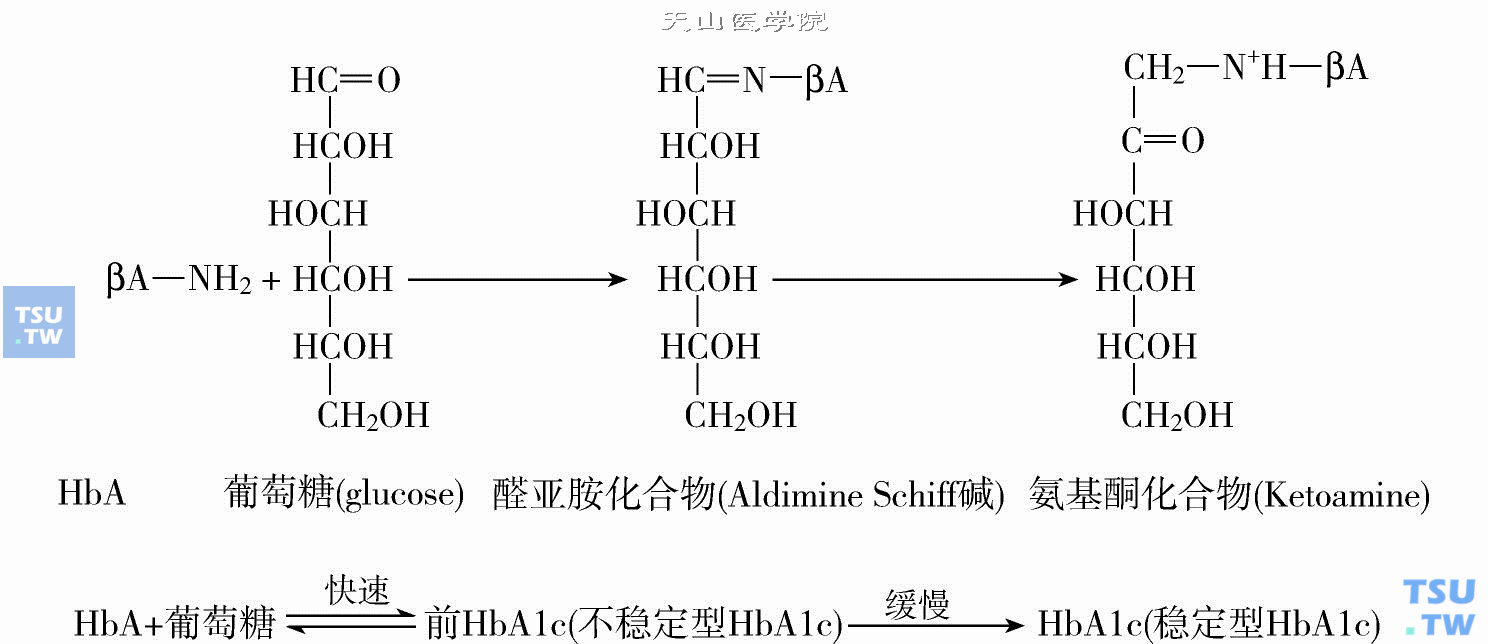
HbA1c的生物合成(引自Bunn HF,1979)
第一阶段为缩合反应,在红细胞中葡萄糖的醛基首先与HbA的β链N端缬氨酸残基发生非酶氨基脱水缩合反应,迅速以共价键形式形成醛亚胺化合物即Schiff碱或称前(Pre-)HbA1c,由于该阶段的反应速度较快且呈可逆还原反应(逆反应较正反应速度慢),故前HbAlc也叫不稳定型HbA1c,其量约占全部反应的10%~20%。
第二阶段为Amadori分子重排反应,醛亚胺化合物经过自动的分子重新排列,通过醛胺基和葡萄糖连接成为酮胺化合物即HbA1c,由于该阶段反应速度缓慢且呈不可逆性故亦称稳定型HbA1c。
由于第一阶段产物是不稳定型HbA1c,其含量变异性较大,时多时少,必然直接影响所测定的结果导致测定值的误差。故从测定方法学要求,应将该部分排除,只测出第二阶段的稳定型HbA1c,才能反映数值的真实性。但有些方法譬如阳离子交换树脂法,如果不预先将血标本中的不稳定型HbA1c进行处理排除,则无法使两者分开。而新近提出的亲和色谱法基于其与稳定型HbA1c呈特异结合的原理,便能排除不稳定型的干扰,只测稳定型HbA1c,成为较理想的方法。HbA1及HbA1c的化学结构:
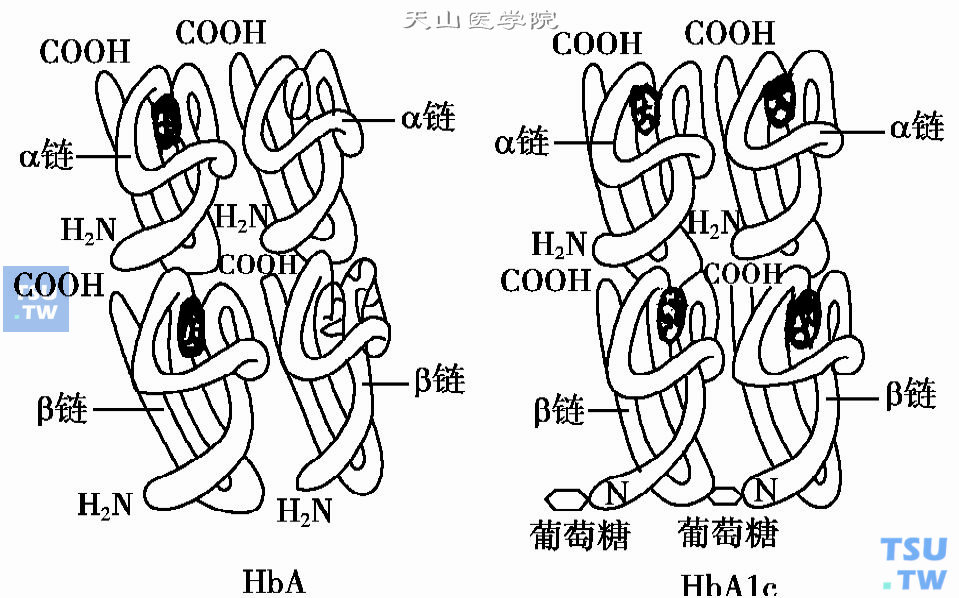
HbA的β链N端与葡萄糖结合所形成的HbA1c的结构模式图(引自长泷重信,1984)
血浆蛋白也可发生非酶糖化反应,糖化血浆蛋白的主要成分是糖化白蛋白,其非酶糖化的机制和血红蛋白相似,唯葡萄糖不一定接在β链的末端氨基上,也可连接在赖氨酸残基的ε氨基上。因白蛋白的半寿期为19~21天,远低于血红蛋白的120天,故糖化白蛋白反映较近期内的血糖平均浓度。
