
维生素K依赖性凝血蛋白的生物合成
参与或调节凝血的血浆凝血因子中有6种因子(包括凝血酶原、FⅨ、FⅩ、FⅦ、PC、PS)的生物合成需维生素K。这些因子氨基(N)端最初45个氨基酸中含10~12个γ-羧基谷氨酸(γ- Gla)(图1),使依赖维生素K的凝血因子能在Ca2+存在条件下在膜表面进行组装而发挥生理功能;因此,这些因子是一种独特的钙结合蛋白,在蛋白合成过程中先以前体蛋白形成出现,含前肽和信号肽。前肽位于信号肽与成熟蛋白N端之间。信号肽能与信号识别小体相结合后,指导蛋白移位到内浆网腔,然后信号肽去除,暴露出前肽(图2)。位于内浆网膜、结合着的γ羟化酶在还原型维生素K、氧分子、二氧化碳及底物(Gla)存在时,将羧化过程与维生素K环氧化过程耦联,使前肽中特殊部位的Gla选择地γ羧化成相应的γ- Gla。前肽在蛋白分子转入高尔基复合体中被去除。

图2 维生素K依赖性凝血因子的γ-羧基谷氨酸富含区的氨基酸顺序(采用单字母氨基酸码,γ代表γ-羧基谷氨酸)
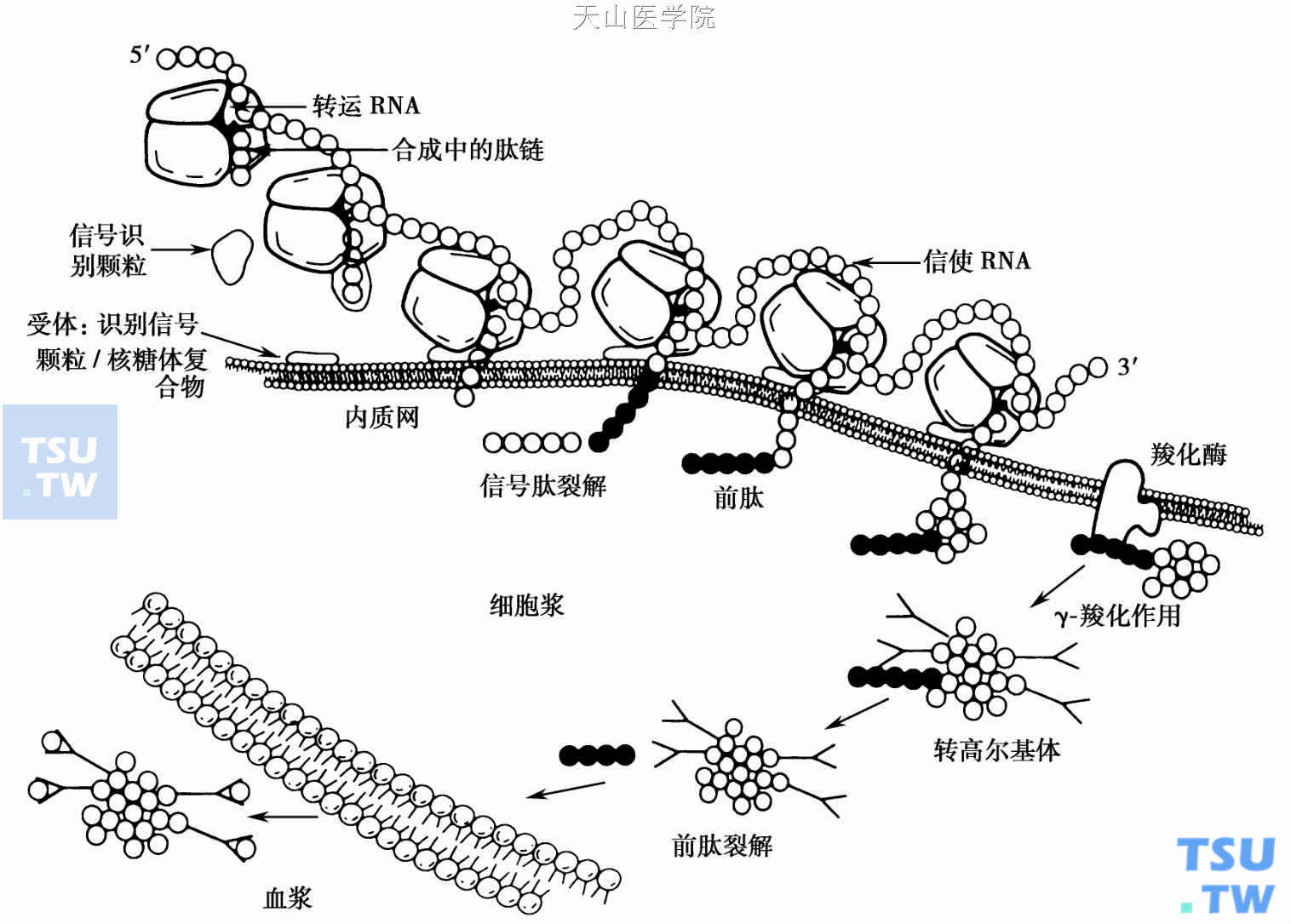
图2 维生素K依赖性凝血因子的生物合成
前肽上的γ-羧化识别位部位(γ- carboxylation recognition site,γ- crs)能引导合成肽按识别系列选择邻近识别部位的Gla加羧基(图3)。此外前肽的另一功能为抑制未成熟蛋白的活化及抑制未成熟蛋白与膜结合。
采用亲和色谱法从FⅨ前肽己纯化得到羧化酶。依赖维生素K的羧化酶(Mr 94 000)是单链糖蛋白含有758个氨基酸的信号肽。昆虫细胞中已能表达此酶,但无活性。
前肽引导的γ羧化作用的识别序列在依赖维生素K的凝血因子中高度同源(图4)。实验表明,全长的前肽才能完全羧化(Km 3μM)。γ-羧化识别部位缺如、点突变或前肽删除则羧化不完全。γ-羧化反应完成后,蛋白被输送到高尔基复合体。高尔基复合体内持续表达的PACE(也称Fiurin)蛋白酶将前肽-1和+1位裂解;去除前肽后的蛋白进一步加工、修饰而成成熟因子。
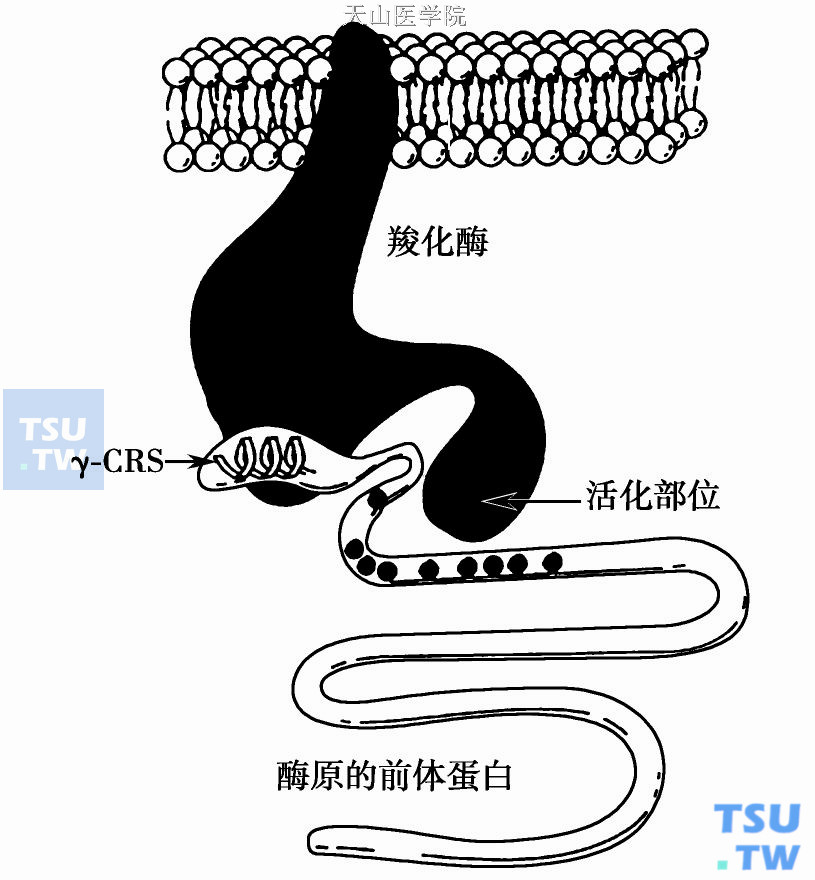
图3 维生素K依赖性凝血因子前体蛋白前肽上的γ-羧化识别部位(γ- CRS)
转录后的β-羟化作用
位于蛋白C(PC)、FⅨ、FⅩ、FⅦ的N端EGF区的红-β-羟天冬氨酸(erythro-β- hydroxyaspartic acid)以及PS中存在的红-β-羟天门冬酰胺都是一种不寻常的氨基酸,能使这些蛋白具有金属结合的特性。这些氨基酸是在蛋白转录后,天冬氨酸和天冬酰胺经羟化作用而产生。β-羟化作用不依赖维生素K 或γ羧化作用。但是,许多维生素K依赖性蛋白以及补体蛋白(C1r,C1s)、凝血酶调节蛋白、低密度脂蛋白受体,存在转录后的这一过程。蛋白EGF区含半胱- X-天冬- X- X- X- X-苯丙/酪- X-半胱- X-半胱序列和转录后的β羟化,需要2-酮戊二酸(2- ketoglutarate)和Fe2+。任何双氧酶(dioxygenases为2-酮戊二酸依赖性酶)抑制剂都能阻断β-羟化过程。

图4 维生素K依赖性凝血因子前肽功能区氨基酸顺序的比较
血管性血友病因子的多聚作用
黏附蛋白von Willebrand因子(vWF)又名血管性血友病因子或FⅧ相关抗原(ⅧR:Ag),是一种异源性血浆多聚体糖蛋白,在内皮细胞和巨核细胞中合成。vWF介导致栓表面与血小板的黏附并引起血小板聚集反应,以及参与血流高切应力下的止血反应。小多聚体vWF是FⅧ:C的载体蛋白。最初翻译产物前肽- vWF单体是单一的多肽链,含信号肽、前肽(很大的前肽Mr 80 000)和成熟vWF(Mr 225 000),在内浆网中以其C-末端的二硫键将两个亚单位联在一起组成二聚体。每一成熟vWF二聚体两侧由邻近N端的二硫键相联。在高尔基复合体中组成多聚体。前肽对多聚体的蛋白二硫键形成起识别作用,并引导vWF到达韦-帕小体(weibelpalade)。vWF高分子量多聚体形成后,作为蛋白生物合成的最后一步为前肽裂解。vWF是一系列多聚体蛋白,分子量自0. 5(二聚体)至2000万(多聚体),亦是唯一由细胞分泌并贮存于细胞内的一种黏附蛋白。电镜下vWF分子呈丝状构型,贮存颗粒中的vWF呈管状结构。巨核细胞合成的vWF贮存于α颗粒中,内皮细胞合成的vWF贮存于韦-帕小体中,胞内贮存为一种调控途径,所组成的vWF库在止血中有重要生理功能。
内皮细胞分泌vWF有两种途径:①持续性分泌途径,在正常情况下细胞未受刺激,合成与分泌耦联,分泌出的是合成的小分子二聚体或小分子多聚体。②受调控的分泌(受刺释放)途径即内皮细胞在某些刺激下脱颗粒导致分泌vWF大分子多聚体。这一分泌过程由促释放素所启动,依赖微管(细胞骨架)而完成。促释放素包括凝血酶、纤维蛋白、组织胺及补体终末期蛋白C5b- 9等。内皮细胞两种途径分泌出的vWF分子大小及生物活性不同。新合成的vWF基本上是通过持续分泌而进入循环(又称管腔分泌)或渗入内膜下基质(腔下分泌)。腔下分泌只发生在动脉、小动脉、大静脉及受刺激的毛细血管基底膜。大多数小多聚体被持续分泌,仅5%贮存于内皮细胞中。小多聚体用于携带FⅧ:C,而大多聚体只是在血管受损部位由内皮或受凝血酶刺激后的血小板(受调控分泌)所释放。释放的大多聚体vWF只在血中短暂存在以促进血小板止血栓形成,并经过有限的蛋白溶解作用在血浆中多聚体裂解、体积变小,故正常血浆中只有少量大分子多聚体。血浆vWF总量的20%贮存于血小板。在骨髓巨核细胞所衍生的血小板的α颗粒中贮存有大聚体,其分泌为受调控式分泌,在正常情况下不进入血流。
血浆中的vWF被血浆中vWF裂解酶(vWF-cleaving protease,vWF- CP)进一步水解成176kDa 和140kDa小分子片段。血栓性血小板减少性紫癜(TTP)患者血浆中vWF- CP缺乏,致使超大分子量vWF多聚体增多。后者促进血小板在高剪切力血管中黏附和聚集反应并因而致病。除TTP之外许多生理和病理状态下存在vWF- CP活性降低,例如新生儿、妊娠中后期妇女及老年人、肿瘤患者等。
(潘家绮)
