
纤维蛋白原及FXⅢ在全血、血浆蛋白、血管壁的互相反应的最后一步中参与止血栓的形成。血浆纤维蛋白原被凝血瀑布反应中最终生成的酶,α凝血酶,转化成不溶性纤维蛋白多聚体。凝血酶也活化血浆转谷氨酰胺酶原(FXⅢ)成FXⅢa。FXⅢa使纤维蛋白多聚体交联,并使其他血浆蛋白能结合在纤维蛋白网络中,包括纤溶系组成及许多黏附蛋白,如纤维连接蛋白、凝血酶敏感蛋白(thrombospondin)、血管性血友病因子(vWF)。这些表面结合蛋白干扰纤维蛋白生成、交联及溶解。纤维蛋白(原)特异地整合膜蛋白结合位点对血小板正常聚集功能十分重要。无纤维蛋白原血症或异常纤维蛋白原血症患者的出血素质能充分说明纤维蛋白原的生理重要性。某些纤维蛋白原功能异常伴血栓前状态。
纤维蛋白原
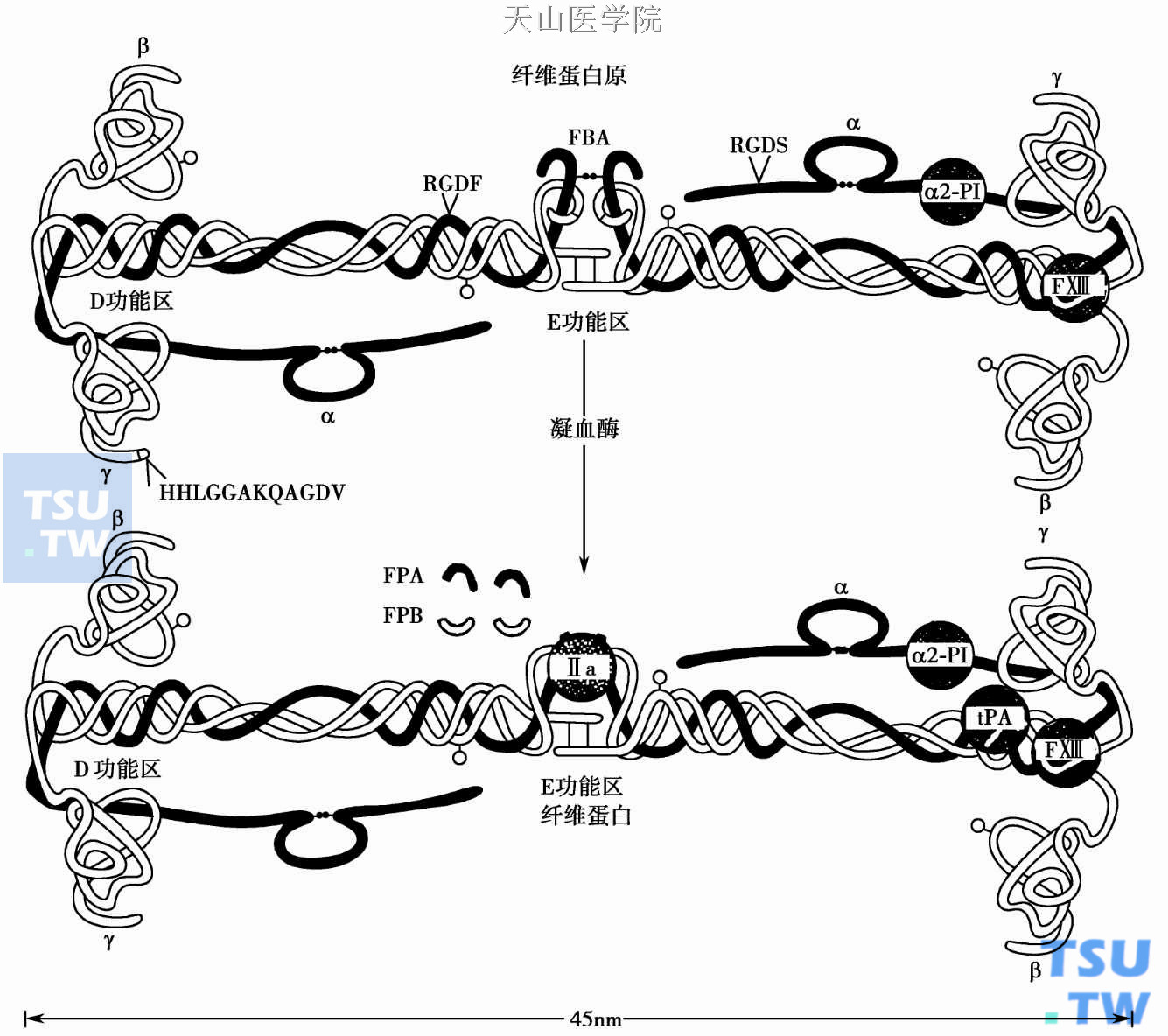
人类纤维蛋白原(fibrinogen或称为凝血因子Ⅰ,Mr 340 000)是一种大分子糖蛋白,存在于血浆(7μm,2~4g/L)和血小板α颗粒中。纤维蛋白原是由3条对称分布的肽链组成的二聚体。肽链名Aα (Mr 665 000)、Bβ(Mr 52 000)和γ链(Mr 46 500)。α凝血酶分别裂解Aα、Bβ链N端的精氨酸16-甘氨酸17、精氨酸14-甘氨酸15键,产生纤维蛋白单体及裂解下的两个小肽片——纤维蛋白肽A(FPA)和纤维蛋白肽B(FPB)。
纤维蛋白原和纤维蛋白的结构示意图
纤维蛋白原中央区为E区,又称二硫键结区(disulfide knot,N- DASK),由以二硫键相联的6条肽键的氨基(N)端组成;外周区(名D区)由3条肽链呈螺旋状盘绕及其球形羧基(C)端所组成。电镜下呈三球状(D- E- D)结构。在肝合成的纤维蛋白原的3条肽键,由不同的基因编码。编码基因位于4号染色体(4q12~q32),全长50kb。α链基因5. 5kb编码2. 2kb mRNA,β链基因8kb编码1. 9kb mRNA,γ链基因8. 5kb编码1. 6kb mRNA。3条基因及其上游序列都高度同源,提示它们复制自同一祖先基因且有相同的调控元件来协调肽键的合成。β链的天冬酰胺364和γ链天冬酰胺52有N-联结的糖基。
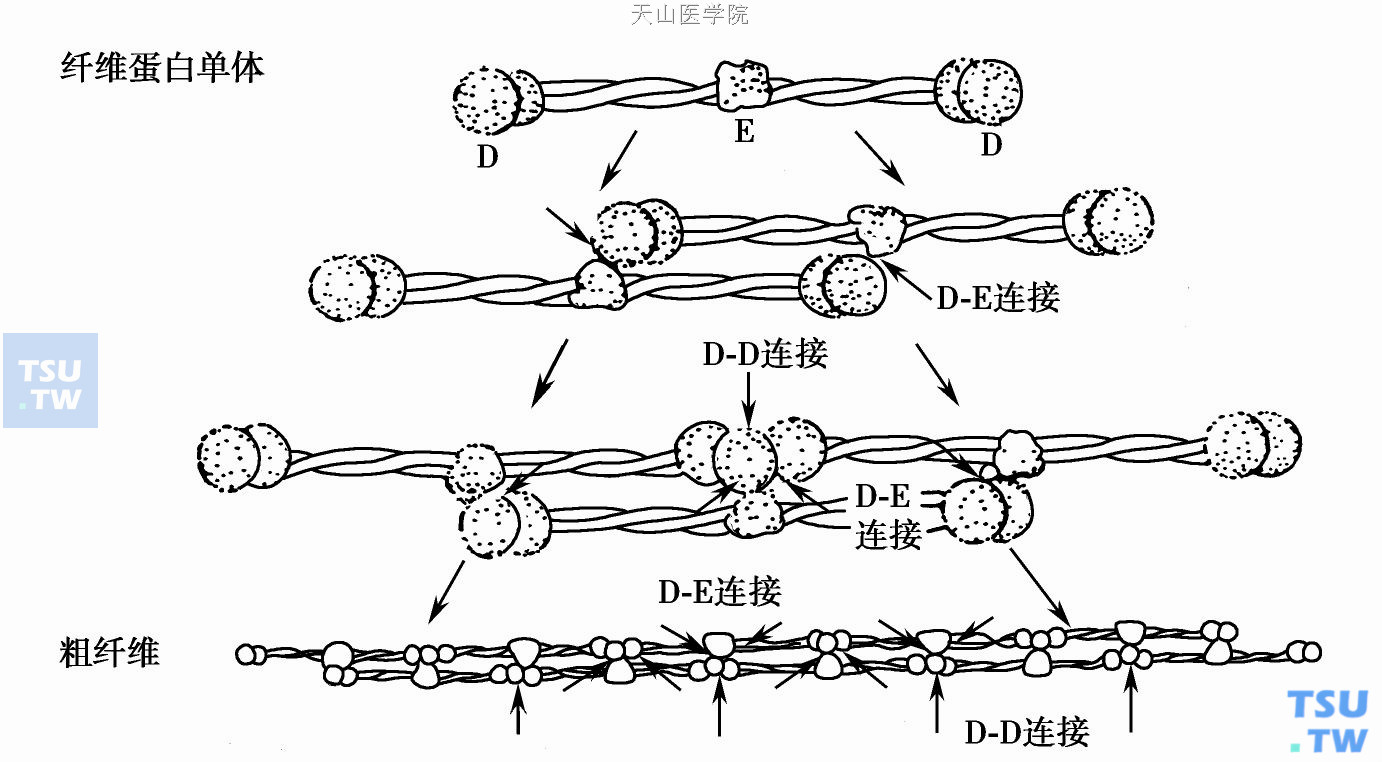
凝血酶和纤维蛋白原的中央区相结合并裂解FPA、FPB,产生纤维蛋白单体。肽键裂解后暴露出的中央区(E)结合部位能与其他纤维蛋白单体分子羧基(C)端(D区)非共价互补(E- D)结合,形成两股有一半重叠(半交叉)构型的原(初始)纤维丝。原纤维丝(E- D)再通过与邻近的第三个分子C端的D- D尾结合,聚集成粗纤维。半交叉的纤维蛋白单体在FXⅢa作用下转谷氨酰胺基而产生共价交叉联结。纤维蛋白存在高、低引力的钙结合位点;钙能增加去FPB小肽后的纤维横向增长。
纤维蛋白粗纤维形成图
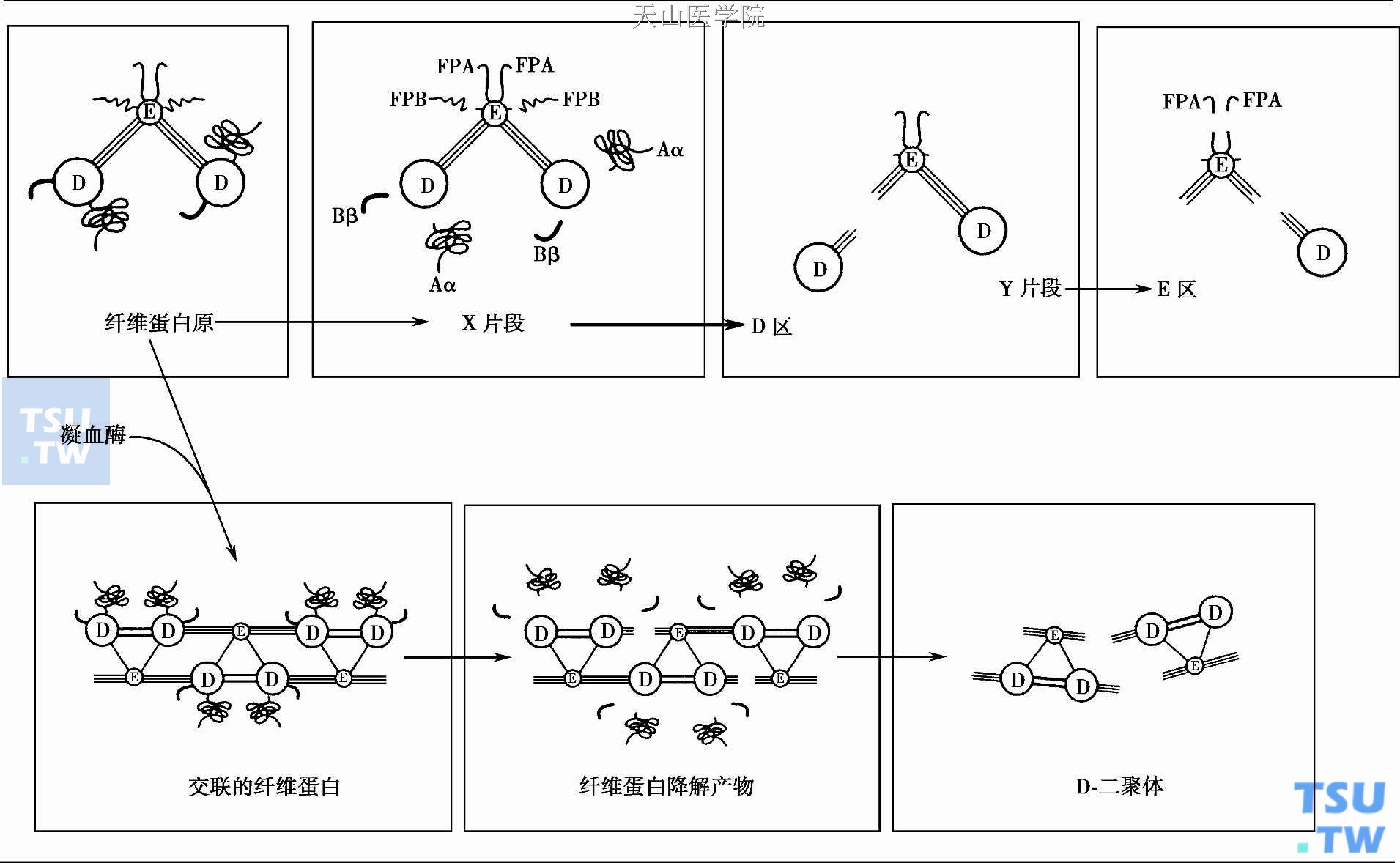
纤溶酶消化纤维蛋白和纤维蛋白原的过程是通过对许多精氨酸、赖氨酸键程序性裂解,产生一系列降解产物(称FDP)。纤溶酶消化纤维蛋白的程序为最初裂解Aα和Bβ1-42片段,产生X碎片(250kDa)。后者仍能形成弱凝块;继之纤溶酶自X碎片裂解下D碎片(100kDa)而产生Y碎片(150kDa);又将Y裂解成另一D和E(50kDa)碎片。下图所示纤溶酶的裂解部位。纤溶酶消化纤维蛋白与上述消化纤维蛋白原的不同之处为自Bβ链上释出Bβ15-42肽片,以及自交叉联结的纤维蛋白多聚体交联部位释出D-二聚体。虽然X碎片仍可多聚化成弱的凝块,但Y和D碎片却抑制纤维蛋白多聚作用。因此,FDPs能使凝血酶凝固时间(TT)测试延长并使基于凝血时间测定的纤维蛋白原定量测定产生假性低值;FDP的可沉淀性又能使基于沉淀法测定的纤维蛋白原定量产生假性高值。D-二聚体单抗能检测来自纤维蛋白的碎片。
纤维蛋白原、纤维蛋白的裂解产物
正常健康人纤维蛋白原血浆半衰期3~5天,仅少量消耗于分解代谢。纤维蛋白原是一种急性反应期蛋白,刺激下的合成速率可增加20倍。白介素-6 (interleukin-6,IL-6)和地塞米松可介导纤维蛋白原合成增加。早期研究报道的FDP能增加纤维蛋白原合成,之后认为是由于单核细胞分泌IL-6的刺激作用。
凝血因子XⅢ
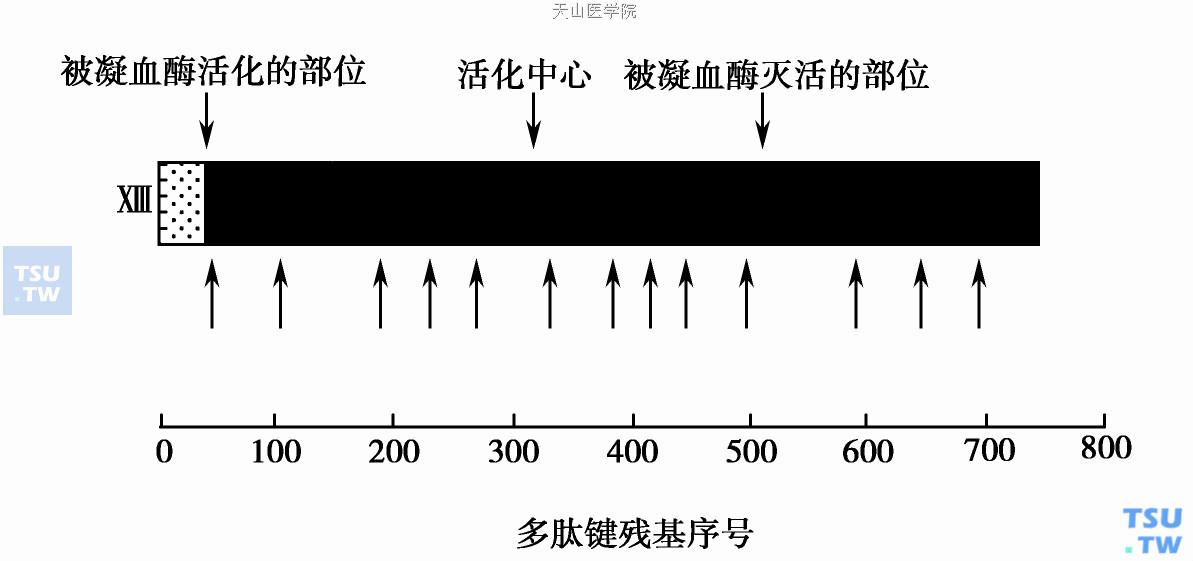
凝血因子XⅢ(FXⅢ)是320kDa糖蛋白,是一种半胱氨酸转谷氨酰胺酶原。FXⅢ有2个不同亚单位,α链(Mr 82 000)和β链(Mr 76 500)各两条(α2β2)组成的四聚体。α链基因169kb,位于6号染色体(6p24- 25),编码3. 8kb mRNA。β链基因28kb,位于1号染色体(1q31- 32),编码2. 3kb mRNA。血浆FXⅢ 2个球形α链与2个丝状β链以非共价键相联,α链含有巯基活化中心起转谷氨酰胺作用;β链起蛋白载体功能并调节酶的功能。血浆四聚体FXⅢ水平0. 07μM(60μg/ml),半衰期9~10天。分布于巨核细胞、血小板α颗粒和单核细胞的FXⅢ细胞内池由2 条α链(β链缺如)所组成;其α链结构与血浆内的α链相同。
FXⅢ活化需凝血酶裂解α链精氨酸37-甘氨酸38键,释出4. 5kDa活化肽片而暴露出半胱氨酸巯基活化中心。下图中箭头表示与多肽链相应的基因外显子-内含子交界区。FXⅢ是一种内-γ碳谷氨酰基-ε氨基-赖氨酸转移酶,能催化邻近的纤维蛋白单体间γ链和α链的赖氨酸和谷氨酸形成ε赖氨酸而产生共价交联的纤维蛋白。酶催化的酰化和脱酰基作用有高度的底物特异性,仅能在底物的少数赖氨酸和谷氨酸基团间进行,底物包括纤维蛋白原、纤维连接蛋白、α2抗纤溶酶、胶原、外连接蛋白、vWF、肌动蛋白、肌球蛋白、因子Ⅴ和凝血酶敏感蛋白。
凝血因子XⅢ亚单位结构示意图
FXⅢα作用的每克分子纤维蛋白有4~6个交联的赖氨酸谷氨酰基:γ链在D区尾对尾交联(C端3个谷氨酰基可供交联);60%Aα链交联。这种α链交联使血块获得稳定以对纤溶酶产生部分抵抗。纤维蛋白与其他蛋白的交联有不同功用,例如:与α2抗纤溶酶交联使α链极性延伸和对控制纤溶酶溶解活性上起作用;α链与黏附蛋白交联可使纤维蛋白止血块与黏附在受损血管内膜下的血小板相结合。
