
已在几百例患者中确定血友病A和B的基因突变,血友病的分子基础是极多样的。人们尚未明确该病的因与果之间的关系,仅在少数突变中明确了血友病如何由存在的基因缺陷引起,这些突变多局限于效应明显的突变,如翻译提前终止,阻碍蛋白活化,或者破坏mRNA剪接等等。对于大多数突变,没有明确解释,一个突变常有多种解释,如蛋白去稳定,不正确折叠,重要区域的结构或功能的混乱等等,通过X线晶体图像和分子模型软件及蛋白质结构预测可能解释某些突变所产生的效应。
目前血友病基因突变研究的结果可以从下列在网上的数据库中得到,血友病A网址:http://europium. csc. mrc. ac. uk/WebPages/Main/main. htm (HAMSTeRS,The Haemophilia A Mutation,Structure,Test,Resource Site),血友病B网址:http://www.kcl. ac.uk/ip/petergreen/haemBdatabase.html(Haemophilia B Mutations Database),二者均能通在下述网址获得:http://archive. uwcm. ac. uk/uwcm/mg/hgmd0. html (Human Gene Mutation Database)。
在FⅧ和FⅨ基因中点突变、缺失、插入和重排/倒位均可见,点突变(单个核苷酸替换)是最常见的基因缺陷,约在90%的患者中存在;缺失是第二位常见的基因缺陷,在5%~10%的患者中存在。除了重型血友病A患者中普遍存在的倒位(这一倒位是内含子22倒位和内含子1倒位)外,插入和重排/倒位在血友病人群中很少见。
CpG烟酰胺腺嘌呤二核苷酸突变
在整个人类基因组中烟酰胺腺嘌呤二核苷酸基因序列CpG中胞嘧啶易发生甲基化;一旦发生甲基化,甲基胞嘧啶残基可能发生脱氨基作用,形成脱氧胸腺嘧啶苷。该种残基属于DNA中含有的核苷酸,不能被体细胞DNA修复机制识别为异常改变。因此,DNA复制产生的新链中原来的GC序列被AC替代。上述过程是整个人基因组发生突变最常见的原因。编码精氨酸的六个遗传密码子中的四个密码子含有CG二核苷酸(CGx),因此,精氨酸密码子突变是最常发生的。在血友病A基因研究中大量的报道描述了错义突变和无义突变发生在精氨酸残基上。在已发现的1082个点突变中,460个(42%)影响到精氨酸残基,除其中的9个意外所有的突变发生在精氨酸密码子(CGx)。
错义突变
错义点突变引起的血友病的严重性取决于替换的氨基酸和它的位置。除非突变发生在结构或功能重要区域,半保守氨基酸替换(新氨基酸与正常氨基酸在电荷,疏水性,极性或形状方面相似)与轻型血友病相关。非保守氨基酸替换(新氨基酸与正常氨基酸非常不同),无论突变发生在蛋白质的何处都会发生中型或重型血友病。对于非保守氨基酸替换,结构紊乱/去稳定的幅度更大,因此它们产生更不利的总效应的可能性更大。
错义突变是血友病A中最常见的基因突变形式,所有重型血友病A均属于该类型突变,血友病的严重程度与突变的类型和位置相关。例如Arg372His位置突变影响到FⅧ激活所必需的凝血酶剪切位点,因此该突变发生后虽然血浆中FⅧ抗原水平正常,但FⅧ活性只有3%~5%;Ser2119Tyr突变导致FⅧ与von Willebrand因子结合力下降,以至于FⅧ失去von Willebrand因子保护,FⅧ:C下降至4%~8%。错义突变可以发生在FⅧ基因所有部位,研究已经证实该类突变可以发生在从起始密码子(19)到终止密码子(2332)。目前为止已经报道有386种错义突变,这些突变影响到了所有氨基酸中10种的某一个氨基酸,而且某些密码子尤其是含有CpG烟酰胺腺嘌呤二核苷酸的密码子常常发生多种不同的突变。三种编码Asp542,Arg2209和Arg2304的密码子是目前已知的最多数目的替代物,每一种可有四种不同的氨基酸代替,例如Arg2209可以被Gln,Gly,Leu和Pro四种氨基酸替代。
无义突变
无义突变产生缩短的蛋白质分子,这样的蛋白分子不可能进入血循环,即使它进入循环,缩短的分子可能迅速消失,因此无义突变与重型血友病相关。外显子跳跃是由无义突变所致,而且这也是很严重的缺陷:一个框内跳跃可导致一个蛋白质缺少跳跃的外显子编码的氨基酸,一个框外跳跃可导致框架移位。
此类突变是由于早熟终端密码子(premature termination codon,PTC)代替某个氨基酸密码子所致。多数情况下PTC被认为导致信使RNA的无义介导衰退(nonsense- mediated decay,NMD)并在细胞内迅速破坏。NMD抑制了异常蛋白的聚集,异常蛋白的聚集可以导致对正常同位基因产生支配性负性作用。在已报道的187例无义突变的血友病A患者中,116例(59. 4%)影响精氨酸残基,如同NMD所预示的,所有此类突变的患者均为重型血友病患者。
mRNA剪接位点突变
血友病的不同严重程度与破坏或产生mRNA剪接位点的突变相关,这主要取决于一些正确的转录是否能被加工(轻到中型血友病)或是否存在正确mRNA加工的完全缺失(重型血友病)。剪接位点突变可能导致外显子跳跃,跳跃发生在读码框内还是读码框移位决定突变的结果。
根据是否为经典的CG位点转变点突变可分为两类,CG位点是高突变区,可引起C- T转变或G- A转变。这些转变的机制已明确,它是人类基因突变的一个主要机制。在血友病中约30%的点突变发生在CG位点,其余70%不发生在CG位点,例如,DNA复制时核苷酸的错误加入引起。也可根据点突变是否重现分为两类,重现突变在血友病A和B中都存在,且主要出现在CG位点。这些位点的高突变性是它们的转变重现的基础,这些通常由在连续的代内增殖所致,这一观点的证据可包括在所有受累基因中局限的区域分布和相同的多态性单倍型,与区域分布无关,Founder突变典型地引起轻/中型血友病。
研究52例血友病A剪切错误突变,发现其中14例受累的是位于5'末端和3'末端内含子中一成不变的GT剪切供体和AG剪切受体位点,此类突变通常导致重型血友病。另有学者研究在10例患者中发现了新的剪切供体和受体位点,其中6例发生在第11外显子,是由于一个CTG>CTT核苷改变产生了一个新的受体剪切位点,导致第11外显子3'末端一个34bp的片段缺失,此类突变导致轻型血友病。目前报道第1内含子A>G改变激活了一个隐藏的剪切位点,导致了新的外显子的转录,此类突变导致重型血友病。
基因片段的缺失和插入突变
FⅧ和FⅨ基因缺失包括全基因缺失,在5'端、3'端或基因内的部分基因缺失,和一个到几个碱基对的小缺失。缺失并非聚集于基因的某特定区域,而是随机分布。缺失可破坏基因功能,蛋白质某一部分缺失,或致读码框移位,所有这些都是极其有害的,因此缺失通常与重型血友病相关。人们已发现在FⅧ和FⅨ基因中的插入突变,如同缺失突变一样,可以是全部或小到一个或几个核苷酸的插入,而且基因功能或基因产物可受到很不利的影响。
目前已报道约105个大片段(一般大于200bp)缺失,其中已发现2例患者发生整个Ⅷ基因缺失(210kb),超过一半的患者发生一个以上的外显子缺失,其余的为单个外显子缺失。目前研究认为基因上散布的重复元件LINE和Alu可能在此类突变中就有重要作用。Water等详尽地研究了大于20. 7kb的大片段缺失突变,发现此类突变是由于起始LINE- 1元件插入FⅧ基因的第20内含子同时与第14内含子的LINE- 1侧序列重组所致,从而导致了第15~20外显子的缺失。另外近来报道一个包含第25外显子约23kb大片段缺失是由于非对等的同源Alu介导的基因重组所致,此类突变是由于第24和25内含子之间的Alu元件重组所致。现在研究显示FⅧ基因是一类Alu丰富的基因,共有52个Alu重复序列在FⅧ工作框架内,每个约3. 8kb,这些序列的重组导致了多种无特征性的FⅧ缺失突变。目前仅报道三个大片段插入突变,L- 1和Alu重复元件整合到FⅧ第14外显子所致。大片段的插入突变和缺失突变导致的血友病均为重型。
小片段基因缺失和插入突变是指小于200bp的片段基因缺失和插入突变。目前FⅧ基因已发现59种小片段插入突变和143种缺失突变,片段大小从1bp到约250bp。单个核苷酸的缺失或插入突变在该类突变中占主导地位。腺嘌呤核苷(A)残基突变是FⅧcDNA最常见的,发生率远远高于其他核苷酸残基,在FⅧcDNA中共有12个A6序列,而只有3 个T6序列,1个G6序列,而无C6序列。在59个插入突变中的32个和143个缺失突变中的31个是A改变所致,其中在FⅧcDNA的1191-4、1439-41和1588- 90是突变发生频率最高的,此类突变是由于DNA复制过程中DNA聚合酶滑脱所致。多数此类突变导致重型血友病,小部分突变是中型血友病。
基因重排/倒位突变
现已明确两个不同的染色体内部基因的倒位所致突变,两者皆为FⅧ内含子中的同源重组序列,此类重组主要发生在男性减数分裂过程中,此时X染色体未配对。
内含子22倒位由内含子22h- 1和内含子22h- 2(近端)或内含子22h- 3(远端)之间在减数分裂时的同源重组引起(图A)。同源排列后,重组导致FⅧ基因的5'端与端粒DNA并排,而适宜的基因外同源序列与FⅧ基因的3'端并排(图B)。在恢复线性化的染色体中,在内含子22h- 1和内含子22h- 2或内含子22h- 3之间的序列发生倒位(图C)。其中与内含子22h- 3的重组更为普遍,也被称为远端倒位(88%倒位),同时其中也包含内含子22h- 2代表的近端倒位(16%倒位)。FⅧ基因被完全打乱:内含子1~22被从它们正常的位置移开,而且它们的方向发生倒转。突变导致基因完全失活,从而形成重型血友病,在全世界范围内约45%的重型血友病由此种突变引起。
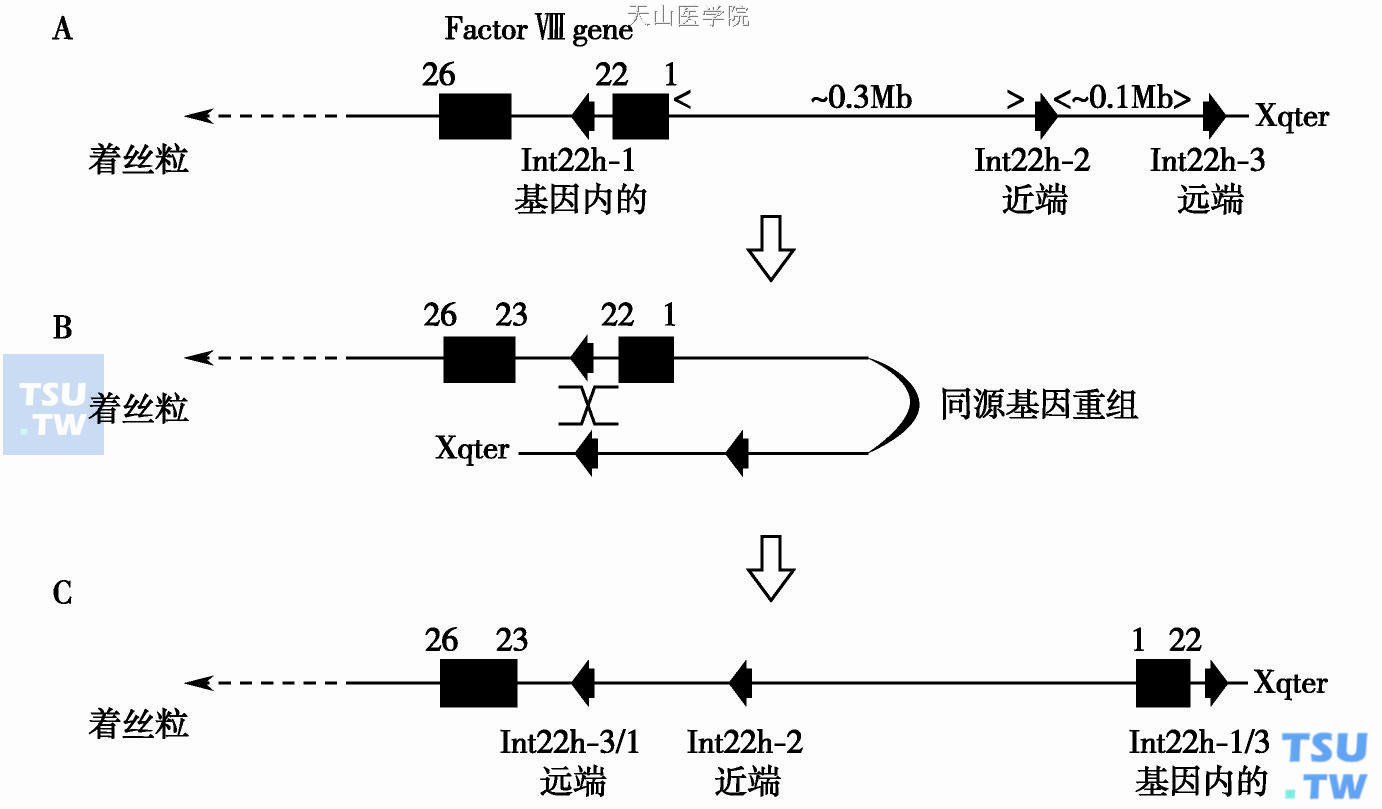
图 FⅧ内含子22倒位
A.正常的FⅧ基因和Int22h同源基因的构型;B.在Int22h基因内部同源基因的远端或近端发生调整,发生同源基因重组;C.从外显子1到外显子22反转,因此,FⅧ的转录过程被打断,不能产生正常的FⅧ
内含子22倒位主要是精子发生而非卵子发生时期DNA复制的一个错误:在男性中,减数分裂时没有第二条X染色体的同源排列(男性只有一条X染色体)将有利于染色体在可能的位置出现染色体内部的排列。因此,在血友病A家族中,内含子22倒位是病因性基因缺陷,这一缺陷可经常表现为来源于一个未受累的男性亲属,而且在散发的病例中,这一缺陷常来自患者的外祖父。远端倒位较近端倒位更常见,这是因为FⅧ基因和远端的内含子22h- 3同源序列之间较大的基因距离有利于排列发生所需基因环的形成。
内含子22倒位用Southern blot分析方法(由正常基因获得的DNA断裂方式不同于由倒位基因获得的断裂方式,而且远端和近端倒位的断裂方式也不同)诊断。这种方法现在已由LD- PCR方法取代。这一方法可检测正常和倒位基因;但是它不能区分近端倒位和远端倒位。虽然大多数内含子22倒位涉及内含子22h- 1和2/3,非典型的倒位也有报道。对一种非典型倒位的详细分析发现了在FⅧ基因和内含子22h- 2之间第三种缩短的内含子22h的拷贝,而且它参与倒位。这些非典型倒位应用Southern blot检测出异常的DNA断裂方式。它们不能用LD- PCR检测出来。
FⅧ基因内含子1中的1. 0kb区域(内含子1h)的另外一个拷贝位于F8基因远端约140kb处这个基因外的区域被称为内含子1h- 2。内含子1h- 1和内含子1h- 2之间的同源重组导致F8基因断裂,启动子和外显子1断开,产生重型血友病。到目前为止这种突变仅仅在英国人群中有报道。与其他的血友病突变一样,这种突变可能也是全世界范围内存在的。大约5%的重型血友病由此突变引起。
通过研究血友病基因突变及其产生效应获得的信息,揭示了血友病的微妙的复杂的分子病理机制,同样从多态性及其种族差异获得的信息暗示一种复杂的进化发展。随着人们对血友病基因突变、基因多态性以及分子病理机制的深入研究,正逐渐为血友病的基因治疗铺平道路,相信不久的将来最终实现治愈血友病。
